Características
físico-químicas y fitoplancton de los humedales altoandinos y su relación con
la presencia de Lymnaea Spp. en Frías, Piura, Perú
Physical-chemical
and phytoplankton characteristics of high Andean wetlands and their
relationship with the presence of Lymnaea Spp. in Frías, Piura, Peru
Luciano Rondoy-Infante*; José Mostacero- León2;
Anthony J. De La Cruz- Castillo2
1 Universidad
Nacional de Piura, Urb. Miraflores S/N, Castilla, 20002, Piura, Perú.
2 Facultad de Ciencias
Biológicas, Universidad Nacional de Trujillo, Av. Juan Pablo II s/n – Ciudad
Universitaria, Trujillo, Perú.
*Autor
corresponsal: lrondoy@hotmail.com (L.
Rondoy-Infante).
ID
ORCID de los autores
J.
Mostacero-León:  https://orcid.org/0000-0003-2556-3013
https://orcid.org/0000-0003-2556-3013
A.
De la Cruz-Castillo: https://orcid.org/0000-0002-5409-6146
DOI: http://dx.doi.org/10.17268/manglar.2020.003
RESUMEN
La presente
investigación se avocó a determinar las características físico-químicas y
fitoplancton de los humedales altoandinos y su relación con la presencia de lymnaea
spp. Frías, Piura, Perú. Se exploró la cantidad de caracoles, la
composición del fitoplancton y variables fisicoquímicas en cinco bofedales
altoandinos, localizados a 3090 msnm. Cada tres meses se colectaron muestras de
agua para análisis fisicoquímico, fitoplancton y caracoles Lymnaea spp.
en lugares georreferenciados, manteniendo el mismo recorrido durante el
muestreo y la misma hora en cada salida de campo. Los análisis fisicoquímicos
del agua indican que presentan poco contenido de calcio y nitrato, así mismo el
contenido total de fitoplancton mostró un ligero efecto sobre el crecimiento
poblacional. Se concluye que los cinco bofedales altoandinos presentan una
riqueza de Fitoplancton, representada en 83 especies, distribuidas en 57
géneros y 5 divisiones: Cyanobacteria, Euglenophyta, Bacillariophyta,
Charophyta y Chlorophyta, siendo la más representativa por su número de especies
las Bacillariophyta; de igual manera existe una asociación positiva entre la
temperatura y el contenido de calcio del agua sobre la densidad poblacional de
los caracoles Lymnaea spp.
Palabras
clave:
fitoplancton; humedales altoandinos; caracoles Lymnaea
spp.
ABSTRACT
The present investigation
aimed to determine the physicochemical and phytoplankton characteristics of the
high Andean wetlands and their relationship with the presence of lymnaea
spp. Frías, Piura, Peru. The number of snails, phytoplankton composition
and physicochemical variables in five high Andean bofedales, located at 3090
meters above sea level, were explored. Water samples were collected every three
months for physicochemical analysis, phytoplankton, and snails Lymnaea spp.
in georeferenced places, maintaining the same route during sampling and the
same time at each field trip. Physicochemical analyzes of the water indicate
that they have little calcium and nitrate content, and the total phytoplankton
content showed a slight effect on population growth. It is concluded that the
five high Andean bofedales present a wealth of Phytoplankton, represented in 83
species, distributed in 57 genera and 5 divisions: Cyanobacteria, Euglenophyta,
Bacillariophyta, Charophyta and Chlorophyta, being the most representative for
their number of species the Bacillariophyta; in the same way there is a
positive association between the temperature and the calcium content of the
water on the population density of the snails Lymnaea spp.
Keywords: phytoplankton;
high Andean wetlands; snails Lymnaea spp.
INTRODUCCIÓN
Perú, con un 0,87% del área total del
planeta (1´285,215.60 km2); se constituye hoy, como el cuarto país megadiverso
del mundo; al poseer cerca del 70 % de la biodiversidad mundial; por no mencionar
que cuenta con 28 de los 32 climas del mundo, 84 de las 117 zonas de vida, 186
Zonas geotérmicas, una variada geografía, edafología, geología y ecología,
reflejada en ocho provincias biogeográficas, tres grandes cuencas
Hidrográficas, 12200 lagos y lagunas, 1007 ríos, así como ingentes humedales
tanto costeros como altoandinos (Brack, 1986; Ginocchio y Acero, 2012;
Mostacero et al., 2007).
Considerándose “humedal”, a aquellas zonas
de tierras planas o con declive moderado, cuya superficie se inunda permanente
o según las condiciones climáticas de la zona; dando lugar a un ecosistema
híbrido muy variado. Tal como ocurre en las zonas Altoandinas del Perú, por un
lado, donde destacan, las ciénagas, los esteros, los fangales, las marismas,
los pantanos, las turberas, y en la costa, por otro lado, representados por los
manglares. Capaces de mantener en equilibrio los ecosistemas típicos de estas
zonas; por no mencionar sus múltiples beneficios, entre los que destacan: su
importancia hídrica, el ser fuente de recursos y energía, su capacidad de
retención de sedimentos y protección, el ser un medio de transporte, su
potencial en recreación y ecoturismo y su inconmensurable importancia ecológica
(RAMSAR, 2009; Tabilo, 1999; Cambra, 2002).
En diferentes partes del mundo se han
realizado investigaciones en humedales de gran altitud, involucrando al
fitoplancton y la calidad del agua; de allí que autores como: Sharma y Singh
(2018) estudiaron la diversidad del fitoplancton y la calidad del agua del
humedal “Dodi Tal”, en la India, con más de 3.075 msnm, los que fueron
monitoreados durante noviembre de 2015 a octubre de 2016. Por otro lado, Scott et
al. (2015) en el Altiplano Chileno, realizaron estudios limnológicos y
ecológicos en los humedales “Lirima” (4000 msnm) y “Caya” (3700 msnm).
Salazar-Torres y Vera (2012) examinaron la diversidad y distribución de las
comunidades de microalgas en bofedales del humedal “Huaytire” en Perú, ubicados
entre 3000 a 4500 msnm. Choy y Anaya (2018) estudiaron la química del agua depositada
y su variabilidad espacio temporal en el bofedal del sector Moyobamba (4600 a
4700 msnm). Salvador et al. (2014) en su estudio en la ecoregión Puna
Peruana apuntaron a describir la condición actual de las turberas en cuanto a
su vegetación, características físicas y químicas y estado de perturbación.
En Frías, Piura, Perú, los humedales
Altoandinos se encuentran a una altitud promedio mayor a 3000 msnm, siendo
ecosistemas muy singulares del Norte del Perú; que albergan una importante
biodiversidad, entre ellos, caracoles pulmonados Lymnaea spp., los que
son esenciales para dar continuidad al ciclo biológico del parásito Fasciola
hepatica; siendo importante, por ello abordar el estado de su población y
como esta se asocia con las características fisicoquímicas y fitoplancton del
agua (Scott et al., 2015). De allí que cambios en las variables de su
hábitat pueden intervenir en su ecología y con ello su dinámica reproductiva;
afectan-do su crecimiento, fecundidad y por consiguiente la supervivencia de
este y con ello cortar el ciclo de la Fasciolasis (Sorensen y Minchella, 1998).
Por lo descrito en los párrafos anteriores
y no habiéndose realizado trabajos de esta índole en los humedales de los
“Altos” de Frías, esta investigación planteó determinar las características
físico-químicas y fitoplancton de los humedales Altoandinos y su relación con
la presencia de Lymnaea spp. Frías, Piura, Perú; a fin de contribuir al
conocimiento sobre el fitoplancton y sus variables fisicoquímicas del agua de
estos humedales; toda vez que estos resultados permitirán integrarse en la
búsqueda del control y erradicación de estos caracoles que permiten la
presencia de Fasciolasis en la zona.
MATERIAL Y
MÉTODOS
Área de estudio
El área de estudio se encuentra circunscrita
a la Sub Cuenca del Rio San Pedro- Chipillico – Chira - Pacífico, y que abarca
a los Centros Poblados de: Méjico, Altos de Poclús, Arenales, Pechuquiz y Las
Pircas en Frías – Ayabaca – Piura – Perú. Ubicados entre los 4o 59' y 4o 52' 30
L. S. y entre los 79o 48' 30 y 79o 54' L. O. (Fig. 1); con una superficie de
112,21 km2 y altitudes que van de los 2900 hasta los 3150 msnm (Pizarro, 2016).
Estos ecosistemas son conocidos en el lugar como “bofedales” u “oconales”. Los
suelos alcanzan hasta el 18% de pendiente (Remigio, 2010), cubierta vegetal
densa y almohadillada. Es decir, conformando comunidades de transición entre
“Césped de Jalca” y “Turberas de Distichia”. Ecológicamente corresponde a la
zona de vida bosque húmedo – Montano Tropical (bh – MT). La temperatura mínima
en la zona oscila entre 6,9 a 13 °C, con mínimas absolutas en Alto Poclús entre
1 y 1,8 °C (SENAMHI, 2016).
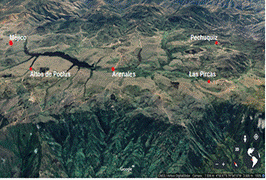
Figura 1. Lugares de muestreo: Bofedales de Méjico, Altos de
Poclús, Arenales, Pechuquiz y Las
Pircas.
Determinación de las características
Físico - químicas
Para cada una de las tres estaciones de
cada humedal se obtuvieron los valores de las variables Físico-químicas del
agua; de allí que para determinar las Características Físicas, fue necesario
realizar la toma de datos climáticos directamente de los cuerpos de agua;
utilizándose para el efecto Termómetro digital VWR® y pHmetro portátil ISOLAB,
con un aproximado de 30 repeticiones, para luego establecer los promedios y
rangos correspondientes.
Para las características químicas se
tomaron 15 muestras de agua en distintos puntos de los 5 humedales Altoandinos
estudiados, separados a una distancia de 100 m unos de otros y convenientemente
georreferenciados con un GPS Garmin eTrex 30. Las muestras previamente
filtradas in situ, fueron preservadas en frascos de vidrio con ácido nítrico y
enviadas al Laboratorio de la Facultad de Ingeniería Química de la Universidad
Nacional de Trujillo, para su análisis por absorción atómica y reporte de los
resultados correspondientes.
Muestreo del fitoplancton
Se realizó filtrando 40 litros de agua
libre, obtenida a una distancia de entre 20 a 80 centímetros del borde de cada
cuerpo de agua, utilizando para tal fin una red de fitoplancton con malla de 10
micrómetros, procediéndose a la fijación de las muestras así tomadas, con
formaldehido al 4%. Para el estudio cuantitativo se tomaron las muestras
directamente utilizando un vaso de precipitación y transfiriéndose de inmediato
150 ml del agua a frascos de vidrio ámbar agregándole formaldehído al 4%.
Análisis cualitativo del
Fitoplancton
Para este análisis, se tomó 1 ml de muestra
previamente homogeneizada y se trasvasó a la cámara de Sedgewick-Rafter para su
lectura en un microscopio invertido, de acuerdo a las normas APHA (2012). En el
estudio se consideró cada individuo algal como una unidad (unicelular, colonia
o filamento). Los resultados de abundancia del fitoplancton fueron expresados
como individuos/litro. Durante el análisis cuantitativo de las algas se tomaron
fotografías con cámara digital adaptada al microscopio invertido.
Colección de los caracoles
Lymnaea spp.
La colección se realizó en forma manual y
de manera exhaustiva e incluyó los materiales presentes en el cuadrante, como
hojas, plantas, piedras, suelo hasta una profundidad de 10 cm, sin estar
condicionada al tiempo de búsqueda. Después de colectados se lavaron y
colocados en un recipiente de plástico rotulado con el nombre del lugar donde
se tomó la muestra, al que se le colocó agua del lugar; siendo transportados al
Laboratorio de Parasitología en cajas térmicas. Los caracoles provenientes de
cada lugar de muestreo fueron contados para determinar la densidad poblacional.
RESULTADOS
Y DISCUSIÓN
En la tabla 1 se presentan
los resultados de los valores promedio y desviación estándar de las
características físico - químicas del agua de los humedales altoandinos en
Frías, Piura, Perú, 2018-2019. En cuanto a las características Físicas, la
temperatura del agua osciló entre 10,99 y 21,98 °C, y el pH fluctuó entre 5,79
y 7,12.
En lo que respecta a las
químicas, el contenido de Calcio varió desde 5,79 hasta 8,5 mg L-1;
y en lo concerniente a Nitratos, su contenido varió de 0.52 hasta 3,13 mg L-1.
Farahnak et al. (2006) establecieron que la temperatura del agua y el pH
favorecieron la emergencia de cercarías desde el caracol, afectando así la
población, por la muerte de los cara-coles parasitados. Se ha encontrado que en
el verano la población de caracoles Lymnaea es más alta que en el otoño
e invierno (Karimi et al., 2004), sin embargo, Prepelitchi et al.
(2011) reportan lo contrario, una menor abundancia en el verano,
incrementándose en el otoño, alcanzando el pico durante el invierno. Según los
resultados obtenidos en el presente estudio, la temperatura del agua de los
bofedales influyó en la dinámica poblacional de los caracoles Lymnaea spp.,
encontrándose que la temperatura entre 18 a 21 °C está asociada a un mayor
crecimiento poblacional de estos gasterópodos; asociadas a la estación de
verano (diciembre-marzo).
Tabla 1
Valores
promedio y desviación estándar de las características físico - químicas del
agua de los humedales altoandinos
|
|
"Méjico"
|
"Alto
Poclús"
|
"Arenales"
|
"Pechuquiz"
|
"Las
Pircas"
|
|
Mes
|
Temperatura
del agua (°C)
|
|
Setiembre
|
15,13 ± 0,65
|
19,4 ± 0,17
|
16,43 ± 0,65
|
19,23 ± 0,89
|
20,9 ± 1,08
|
|
Diciembre
|
19,73 ± 0,58
|
17,9 ± 0,70
|
18,6 ± 0,53
|
18,57 ± 0,64
|
19,87 ± 0,83
|
|
Marzo
|
13,77 ± 1,05
|
12,9 ± 1,30
|
12,5 ± 1,51
|
13,07 ± 1,31
|
12,87 ± 1,32
|
|
pH
|
|
Setiembre
|
6,21 ± 0,30
|
5,96 ± 0,21
|
6,31 ± 0,12
|
6,20 ± 0,35
|
6,50 ± 0,04
|
|
Diciembre
|
6,41 ± 0,10
|
5,84 ± 0,05
|
6,16 ± 0,13
|
6,16 ± 0,21
|
6,54 ± 0,29
|
|
Marzo
|
6,63 ± 0,23
|
6,15 ± 0,16
|
6,48 ± 0,24
|
6,55 ± 0,31
|
6,81 ± 0,31
|
|
Calcio
(mg/L)
|
|
Setiembre
|
7,23 ± 1,27
|
3,07 ± 0,84
|
4,11 ± 0,05
|
2,80 ± 0,61
|
5,15 ± 2,40
|
|
Diciembre
|
6,93 ± 1,30
|
3,15 ± 0,71
|
4,26 ± 0,25
|
2,79 ± 0,52
|
4,87 ± 1,94
|
|
Marzo
|
6,98 ± 1,96
|
3,49 ± 0,34
|
4,18 ± 0,21
|
2,87 ± 0,48
|
4,85 ± 2,05
|
|
Nitrato
(mg/L)
|
|
Setiembre
|
1,21 ± 0,66
|
1,40 ± 0,57
|
2,36 ± 0,77
|
1,41 ± 0,46
|
1,53 ± 0,33
|
|
Diciembre
|
1,22 ± 0,60
|
1,35 ± 0,61
|
2,32 ± 0,87
|
1,43 ± 0,52
|
1,63 ± 0,34
|
|
Marzo
|
1,17 ± 0,65
|
1,34 ± 0,57
|
2,28 ± 0,68
|
1,39 ± 0,51
|
1,58 ± 0,25
|
En las Figuras 2 y 3 se
puede apreciar la influencia entre la temperatura del agua y el contenido de
calcio sobre la densidad poblacional de Lymnaea spp.; apreciándose un
incremento en la población de estos especímenes, cuando la temperatura del agua
va alcanzando valores mayores a 19 °C, no sucediendo lo mismo con los niveles
de pH.
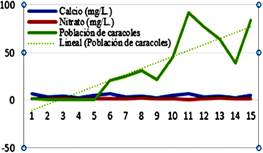
Figura 2. Relación entre
la temperatura y el pH del agua con la densidad poblacional de caracoles Lymnaea
spp.
Por otro lado, se aprecia
un efecto positivo sobre el crecimiento poblacional de estos caracoles por
influencia del calcio, a pesar de que este mineral se encuentra presente en
pequeñas concentraciones. En lo que respecta al nitrato no se observó influencia
alguna sobre la densidad poblacional de estos gasterópodos pulmonados.
Prepelitchi et al.
(2011) encontraron que Lymnaea columella fue muy activo en su hábitat
durante todo el año con valores de pH entre 5,5 – 7,8, mientras que Islam et
al. (2012) encontraron que Lymnaea auricularia y Lymnaea luteola
vivieron en aguas con un pH de 5,7 – 5,9 y a temperaturas con un rango de 24,8
– 27,3 oC. En todos los bofedales evaluados situados a altitudes
aproximadas de 3 090 msnm sus aguas fueron ácidas, difiriendo de lo encontrado
por Sulca et al. (2019) quienes reportan pH ligeramente neutro en
bofedales a una altitud de 4440 a 4556 msnm, y por lo obtenido por Scott et
al. (2015), quienes registraron valores de 8,3 y 8,7. Existe concordancia
con el estudio realizado por Benavides et al. (2013) en turberas, los
que obtuvieron valores entre 4,9 a 5,8 y por lo realizado por Salvador et al.
(2014) en turberas de una ecorregión de Puna en el Perú, informando de valores
promedio de 6,3 y 6,5. El hecho de que las aguas fueron más ácidas en Alto de
Poclús y menos ácidas en Las Pircas, puede estar asociado a las cantidades de las
comunidades de microalgas, quienes al consumir el CO2 que se
encuentra en el agua para llevar a cabo la fotosíntesis, disminuye la probabilidad
de la formación del ácido carbónico (Almomani et al. 2019; Liu et al.
2019). La temperatura del agua puede haber contribuido a una menor solubilidad
del dióxido de carbono atmosférico, estando establecido que la solubilidad del
CO2 disminuye al aumentar la temperatura (Christmas y
Bassingthwaighte, 2017), coincidiendo con la mayor temperatura registrada en
Las Pircas.
Por otro lado, el hecho de
que el contenido de calcio de las aguas sea muy bajo (Tabla 1, Fig. 4), contrasta
quizás con lo reportado por Scott et al. (2015) quienes en humedales
ubicados a una altitud de 4000 y 3700 msnm; encontraron 60,9 y 261,1 mg L-1
de calcio medido por absorción atómica. Benavides et al. (2013) en
turberas reportan valores de 7,3 a 17,4 mg L-1 para altitudes de
4300 a 4700 msnm. Salvador et al. (2014) hallaron valores entre 35,3 y
48 mg L-1. Esta baja concentración de Calcio (Tabla 1, Fig. 4)
podría estar relacionada con una reducida concentración de sales inorgánicas de
calcio en el agua y al tipo de suelo de la zona; esto es característico de
ecosistemas ombrotróficos donde la fuente principal de agua es de lluvia
(Acero-Rodríguez y León, 2016).
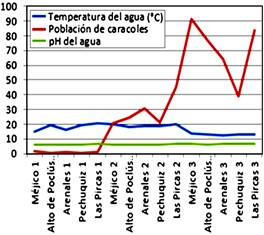
Figura 3. Relación entre
la concentración de calcio y nitrato presente en el agua con la densidad
poblacional de caracoles Lymnaea spp.
Los valores de nitratos
son mayores a encontrados por Corredor et al. (2016) quienes reportan
0,03 a 0,09 mg L-1. Sin embargo, se pueden considerar como valores
medianos. Corredor et al. (2016) citan que los valores bajos de nitratos
en agua se deben a una buena oxigenación de esta, hecho que probablemente no
ocurra. A ello puede sumarse la actividad nitrificante y desnitrificante de los
microorganismos presentes en el agua (Corredor et al. 2016), entre ellas
las microalgas, hay que recordar que el en la fase de oxidación el nitrato re-presenta
la fase más alta en el ciclo del nitrógeno y alcanza normalmente,
concentraciones importan-tes en las etapas finales de la oxidación biológica.
En la Tabla 2 se muestra
la densidad poblacional de los caracoles Lymnaea spp. según el bofedal
evaluado. Se muestra que los bofedales de Méjico y Las Pircas presentaron la
mayor densidad poblacional con una regularidad según la estación del año.
Tabla 2
Densidad poblacional
promedio de caracoles Lymnaea spp. por m2 en los lugares de
muestreo
|
Lugar de Muestreo
|
Setiembre
|
Diciembre
|
Marzo
|
|
Méjico
|
1,67
|
20,67
|
91,33
|
|
Alto
Poclús
|
0,67
|
24,67
|
76,67
|
|
Arenales
|
1
|
30,67
|
64
|
|
Pechuquiz
|
0,33
|
21,33
|
39
|
|
Las Pircas
|
1
|
45,33
|
84
|
Cabe destacar que se ha
establecido que la densidad poblacional no es igual según el hábitat y la
distancia de muestreo teniendo como base la orilla del cuerpo de agua (Utzinger
y Tanner, 2000); concordando con los hallazgos establecidos en el campo, aunque
con diferencias en las cantidades, probablemente afectado por la complejidad
del sustrato de los bofedales evaluados, que puede afectar la ubicación de los
caracoles.
En la Tabla 3 se muestra
la riqueza de fitoplancton encontrado, correspondiente a 83 especies distribuidas
en 57 géneros y 5 Phylum: Bacillariophyta, Charophyta, Chlorophyta,
Cyanobacteria y Euglenophyta. Se ha postulado que una disminución en el
crecimiento algal y la falta de plantas acuáticas podría explicar una baja
población de caracoles (Prepelitchi et al., 2011). Considerando que no se
encontró un efecto significativo en la densidad poblacional de caracoles
asociada a las especies de microalgas encontradas y a su número, es probable
que sean las plantas hidrófitas las que influyan más en el sostenimiento de las
poblaciones de caracoles. La mayor prevalencia de Navicula sp. (86,67%)
de los lugares estudiados, seguido de Ulnaria ulna en 9 lugares (60%) y Gomphonema
sp. en 8 lugares (53,33 %), parecen no influir en la dinámica poblacional
de los Lymnaea spp. En el caso de las algas verdes prevaleció Closterium
sp. hallándosele en 11 de los lugares evaluados, lo que representa el 73,33
%. Según Escobar et al. (2013) esta es una especie cosmopolita en aguas
dulces y habita en aguas ácidas lo que está en correspondencia con los valores
registrados de pH del agua. En cyanobacterias prevalecieron Anabaena sp
en 6 lugares (40%) y Phormidiun sp., en 8 lugares (53,33 %). En
Euglenophytas prevaleció Trachelomonas hispida (46,66%) seguido de Euglena
sp. (26,67%).
La Figura 4 detalla la
asociación entre la cantidad total de fitoplancton y la densidad poblacional de
los caracoles Lymnaea spp., encontrándose una ligera tendencia positiva
entre estas variables.
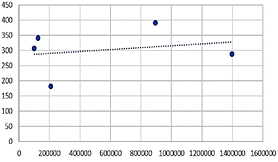
Figura 4.
Asociación
entre la cantidad de fitoplancton y la densidad poblacional de los caracoles Lymnaea
spp.
Tabla 3
Fitoplancton de los
humedales altoandinos y su relación con la presencia de Lymnaea spp.
Frías, Piura, Perú
|
Méjico
|
Alto de
Poclús
|
Arenales
|
Pechuquiz
|
Las Pircas
|
|
|
E1
|
E2
|
E3
|
E1
|
E2
|
E3
|
E1
|
E2
|
E3
|
E1
|
E2
|
E3
|
E1
|
E2
|
E3
|
|
Bacillariophyta
|
7
|
8
|
10
|
2
|
5
|
3
|
9
|
9
|
10
|
11
|
10
|
13
|
5
|
5
|
3
|
|
Achnanthidium minutissimun
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Amphora
sp.
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Cocconeis
placentula
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Craticula
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Cymatopleura
solea
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
|
Cymbella
sp.
|
-
|
X
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
X
|
X
|
-
|
X
|
|
Denticula
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Encyonema
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Epithemia
sorex
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
|
Epithemia
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
X
|
-
|
-
|
X
|
-
|
X
|
-
|
|
Fragilaria
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
-
|
-
|
-
|
-
|
|
Frustulia
sp.
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
|
Gomphonema
acuminatum
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Gomphonema
cf.
gracile
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Gomphonema
sp.
|
-
|
X
|
-
|
X
|
X
|
-
|
X
|
-
|
X
|
X
|
-
|
-
|
-
|
X
|
X
|
|
Gomphonema
subclavatum
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
|
Gomphonema
truncatum
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Gyrosigma
sp.
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Hantzschia
amphioxys
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
X
|
-
|
X
|
-
|
-
|
|
Luticola
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
|
Melosira
varians
|
X
|
X
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
X
|
-
|
X
|
-
|
|
Navicula
sp.
|
X
|
X
|
X
|
-
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
-
|
X
|
|
Nitzschia
cf. Elongata
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Nitzschia
sp.
|
-
|
-
|
X
|
-
|
X
|
-
|
-
|
X
|
X
|
-
|
X
|
X
|
-
|
-
|
-
|
|
Pinnularia
sp.
|
-
|
-
|
-
|
X
|
X
|
X
|
X
|
-
|
X
|
-
|
X
|
X
|
-
|
-
|
-
|
|
Pinnularia
viridis
|
X
|
X
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
X
|
X
|
-
|
-
|
|
Rhopalodia
gibba
|
-
|
-
|
X
|
-
|
X
|
-
|
X
|
-
|
X
|
X
|
-
|
X
|
-
|
X
|
-
|
|
Surirella
sp.
|
X
|
X
|
X
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Ulnaria
ulna
|
X
|
X
|
X
|
-
|
-
|
-
|
X
|
X
|
-
|
X
|
X
|
X
|
-
|
X
|
-
|
|
Charophyta
y Chlorophyta
|
5
|
5
|
1
|
3
|
3
|
17
|
1
|
3
|
12
|
4
|
4
|
3
|
19
|
3
|
3
|
|
Ankistrodesmus
falcatus
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Closterium
kuetzingii
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Closterium
lunula
|
X
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
X
|
-
|
-
|
-
|
-
|
-
|
|
Closterium
sp.
|
X
|
-
|
X
|
-
|
-
|
X
|
-
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
X
|
|
Coelastrum
sp.
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Cosmarium
sp
|
|
X
|
|
|
|
X
|
|
|
X
|
|
|
X
|
X
|
|
|
|
Cosmarium
sp.1
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Cosmarium spp. 2
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Crucigenia
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Desmidium
cf. aptogonum
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Desmidium
sp.
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Desmodesmus
brasiliensis
|
-
|
-
|
-
|
-
|
X
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Desmodesmus
quadricauda
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
|
Desmodesmus
sp.
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Eudorina
sp.
|
|
|
|
|
|
X
|
|
|
|
|
X
|
|
X
|
X
|
|
|
Gloeocystis
sp.
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Micrasterias
laticeps var. laticeps
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Oedogonium
sp.
|
-
|
-
|
-
|
X
|
X
|
X
|
X
|
-
|
-
|
-
|
X
|
-
|
X
|
-
|
-
|
|
Oocystis
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Pandorina
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
|
Pediastrum
boryanum
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Penium
sp.
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Pleurotaenium
cf. ehrenbergii
|
-
|
-
|
-
|
X
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Pleurotaenium
cf. trabecula
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Pseudopediastrum boryanum
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Scenedesmus
cf.
ecornis
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
|
Scenedesmus
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Sorastrum
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Spirogyra
sp.
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Staurastrum
cf.
margaritaceum
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Staurastrum
sp.
|
-
|
X
|
-
|
X
|
-
|
X
|
-
|
X
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Staurastrum
sp.1
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Stauridium
tetras
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
-
|
|
Staurodesmus
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
|
Tetradesmus
lagerheimii
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Tetradesmus
obliquus
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Tetradesmus dimorphus
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Zygnema
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
|
Cyanobacteria
|
1
|
1
|
1
|
0
|
0
|
1
|
1
|
3
|
3
|
2
|
2
|
3
|
2
|
1
|
2
|
|
Anabaena
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
X
|
X
|
X
|
-
|
X
|
-
|
X
|
|
Anabaena
sp.1
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
|
Aphanocapsa
sp.
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Nostoc
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
|
Oscillatoria
princeps
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Oscillatoria
sp
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Phormidium
sp.
|
-
|
X
|
X
|
-
|
-
|
-
|
X
|
X
|
X
|
X
|
X
|
X
|
-
|
-
|
-
|
|
Pseudanabaena
sp.
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
|
Euglenophyta
|
1
|
0
|
1
|
1
|
4
|
2
|
1
|
0
|
5
|
0
|
3
|
1
|
0
|
2
|
0
|
|
Euglena
sp.
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
X
|
-
|
-
|
X
|
-
|
|
Euglena
spirogyra
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Lepocinclis
sp.
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
X
|
-
|
X
|
-
|
-
|
-
|
-
|
|
Phacus
cf. helikoides
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Phacus
sp.
|
-
|
-
|
-
|
X
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Trachelomonas
armata
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
-
|
|
Trachelomonas
hispida
|
X
|
-
|
X
|
-
|
X
|
-
|
X
|
-
|
-
|
-
|
X
|
X
|
-
|
X
|
-
|
|
Trachelomonas
sp.
|
-
|
-
|
-
|
-
|
-
|
X
|
-
|
-
|
X
|
-
|
-
|
-
|
-
|
-
|
-
|
|
TOTAL
DE ESPECIES
|
14
|
14
|
13
|
6
|
12
|
23
|
12
|
15
|
30
|
17
|
19
|
20
|
26
|
11
|
8
|
CONCLUSIONES
Los cinco bofedales
altoandinos de Frías, presentan una riqueza de Fitoplancton, representada en 83
especies, distribuidas en 57 géneros y 5 divisiones: Cyano-bacteria,
Euglenophyta, Bacillariophyta, Charophyta y Chlorophyta, siendo la más
representativa por su número de especies las Bacillariophyta.
Existe una asociación
positiva entre la temperatura y el contenido de calcio del agua sobre la
densidad poblacional de los caracoles Lymnaea spp. y una pobre relación
con el contenido de nitratos y el fitoplancton.
REFERENCIAS
BIBLIOGRÁFICAS
Acero-Rodríguez, M.; León, Y. 2016.
Caracterización de una turbera de Sphagnum L. asociada al bosque Montano en
Mérida, Venezuela. Acta Botanica Venezuelica 39(2): 204-230.
Almomani, F.; Al Ketife, A.; Judd, S.;
Shurair, M.; Bhosale, R.; Znad, H.; Tawalbeh, M. 2019. Impact of CO2
concentration and ambient conditions on microalgal growth and nutrient removal
from wastewater by a photobioreactor. Science of The Total Environment 662:
662-671.
APHA (American Public Health Association).
2012. Standard Methods for the examination of water and Wasterwater. 22nd.
American Public Health Association, American Water Works Association, Water
Environment Federation.
Benavides, J.; Vitt, D.; Wieder, R. 2013.
The influence of climate change on recent peat accumulation patterns of Distichia
muscoides cushion bogs in the high-elevation tropical Andes of Colombia. Journal
of geophysical research: biogeosciences 118: 1627-1635.
Brack, A. 1986. Ecología de un país
complejo. En Gran Geografía del Perú. Vol. II. Edit. ManferJuan Mejía Baca. Barcelona-
España.
Cambra, P. 2002. ¿Réquiem para los
manglares? Rev. Ambiente ecológico N.° 57. Panamá.
Choy, A.; Anaya, M. 2018. Hidroquímica y su
variabilidad espacio temporal en un bofedal altoandino de la Reserva
Paisajística Nor Yauyos Cochas, Perú. IDESIA (Chile) 36(4): 87-97.
Christmas, K.; Bassingthwaighte, J. 2017.
Equations for O2 and CO2 solubilities in saline and plasma: combining
temperature and density dependences. Journal of applied physiology 122(5):
1313-1320.
Corredor, L.; Galarza, G.; Hernandez, E.;
Macariz, A.; Tamayo, P.; Vallejo, L. 2016. Análisis físico y químico en cuatro
ecosistemas acuáticos en el valle del cauca, Colombia. Monografía. Bogotá,
Departamento de Biología, Facultad de Ciencias, Universidad Nacional de
Colombia.
Escobar, M.; Terneus, E.; Yánez, P. 2013. El
plancton como bioindicador de la calidad del agua en zonas agrícolas andinas:
análisis de caso. Qualitas 5(1):17-37.
Farahnak, A.; Vafaie-Darian R.; Mobedi, I.
2006. Faunistic Survey of Cercariae from Fresh Water Snails: Melanopsis spp.
and their Role in Disease Transmission. Iranian J Publ Health 35(4): 70-74.
Ginocchio, L.; Acero, R. 2012. Nueva Ley
Forestal y de Fauna Silvestre: Por un aprovechamiento sostenible, equitativo y
competitivo de los bosques en el Perú. Dirección General Forestal y de Fauna
Silvestre, Ministerio de Agricultura. 122 pp.
Islam, Z.; Alam, M.; Akter, S.; Roy, B.;
Mondal, M. 2012. Distribution Patterns of Vector Snails and Trematode Cercaria
in their Vectors in Some Selected Areas of Mymensingh. J. Environ. Sci. &
Natural Resources 5(2): 37- 46.
Karimi, G.; Derakhshanfar, M.; Paykari, H.
2004. Population Density,Trematodal Infection and Ecology of Lymnaea Snails in
Shadegan, Iran. Arch. Razi Ins 58: 125-129.
Liu, S.; Elvira, P.; Wang, Y.; Wang, W.
2019. Growth and Nutrient Utilization of Green Algae in Batch and
Semicontinuous Autotrophic Cultivation Under High CO2 Concentration. Applied
Biochemistry and Biotechnology 188(3): 836-853.
Mostacero, J.; Mejía, F.; Zelada, W.;
Medina, C. 2007. Biogeografía del Perú. Editorial Asamblea Nacional de
Rectores. Trujillo - Perú.
Pizarro, I. 2016. Diagnóstico de la Meseta
Andina Central. Disponible en: https://docplayer.es/86509953-Ing-isabel-del-pilar-pizarro-cornejo-consultora.html
Prepelitchi, L.; Pietrokovsky, S.; Kleiman,
F.; Rubel, D.; Issia, L.; Moriena, R.; Racioppi, O.; Álvarez, J.;
Wisnivesky-Colli, C. 2011. Population Structure and Dynamics of Lymnaea
columella (Say, 1817) (Gas-tropoda: Lymnaeidae) in Wetlands of Northeastern
Argentina. Zoological Studies 50(2): 164-176.
RAMSAR. 2009. Manual de la Convención de
Ramsar: Guía a la Convención sobre los Humedales (Ramsar, Irán, 1971), 4a.
edición. Secretaría de la Convención de Ramsar, Gland (Suiza).
Remigio, J. 2010. Servicio de consultoría
para ejecutar el estudio de capacidad de uso mayor de la tierra en el
departamento de Piura. Estudio CUM Región Piura. Tercer Informe (FINAL). Piura.
Perú. 151 pp.
Salazar-Torres, G.; Vera, L. 2012. Microalgae
community of the Huaytire wetland, an Andean high-altitude wetland in Peru.
Acta Limnol. Bras 24(3): 285-292.
Salvador, F.; Monerris, J.; Rochefort, L.
2014. Peatlands of the Peruvian Puna ecoregion: types, characteristics and
disturbance. Mires and Peat 15(3): 1–17.
Scott, S.; Dorador, C.; Oyanedel, J.;
Tobar, I.; Hengst, M.; Maya, G.; Harrod, Ch.; Vila, I. 2015. Microbial
diversity and trophic components of two high altitude wetlands of the Chilean
Altiplano. Gayana 79(1): 45-56.
SENAMHI - Servicio Nacional de Meteorología
e Hidrología. 2016. Caracterización y Escenarios Climáticos para la Región
Piura. Integración metodológica de la investigación para incorporar la variable
de cambio climático en los procesos de planificación de las regiones de Piura y
Tumbes en el marco del TACC. PNUD. Perú.
Sharma, R.; Singh, S. 2018. Water quality
and phytoplankton diversity of high-altitude wetland, Dodi Tal of Garhwal
Himalaya, India. Biodiversity International Journal 2(6): 4894-493.
Sorensen, R.; Minchella, D. 1998. Parasite
influences on host life history: Echinostoma revolutum parasitism of Lymnaea
elodes snails. Oecologia 115: 188-95.
Sulca, L.; Franco, P.; Oyague, E. 2019. Caracterización
trófica de dos bofedales de la provincia de Candarave, Región Tacna. Ciencia
& Desarrollo 21(2): 37-49.
Tabilo, V. 1999. El beneficio de los
humedales en América Central: El potencial de los humedales para el desarrollo.
2da edición. Editorial Universidad Nacional Heredia. San José, Costa Rica.
Utzinger, J.; Tanner, M. 2000. Microhabitat
Preferences of Biomphalaria pfeifferi and Lymnaea natalensis in a Natural and a
Man-made Habitat in Southeastern Tanzania. Mem Inst Oswaldo Cruz 95(3):
287-294.