Reguladores
de Crecimiento (BAP y ANA) en la Propagación in vitro de Queñoa (Polylepis
rugulosa Bitter)
Growth
Regulators (BAP and ANA) for in vitro Propagation of Queñoa (Polylepis
rugulosa Bitter)
Carmen Mollohuanca Ccuno1, *; Luis
Mayta Anco1; Roxana Bardales Alvarez2
1 Laboratorio
de Biotecnología Vegetal, Sociedad Minera Cerro Verde, Av. de Las Tradiciones
S/N, Congata, Uchumayo, Arequipa, Perú.
2 Facultad de Agronomía,
Universidad Nacional de San Agustín de Arequipa, Urb. Aurora S/N, Arequipa, Perú.
*
Autor corresponsal: c.mollohuanca@conexxaperu.com
(C. Mollohuanca Ccuno).
ID
ORCID de los autores
C.
Mollohuanca Ccuno:  http://orcid.org/0000-0002-9105-8501
http://orcid.org/0000-0002-9105-8501
L.
Mayta Anco: http://orcid.org/0000-0001-8359-4876
R.
Bardales Alvarez: http://orcid.org/0000-0002-5137-4067
RESUMEN
Dentro del marco del
Programa de forestación e investigación forestal de Sociedad Minera Cerro Verde
se realizó el presente estudio, con la finalidad de establecer un protocolo de
propagación vegetativa mediante la técnica de propagación in vitro de yemas de
queñoa (Polylepis rugulosa Bitter) para su conservación, por ser una
especie forestal en peligro, para lo cual se realizó el cultivo con diferentes
concentraciones de auxina y citoquinina en la fase de multiplicación in vitro,
pasando por la fase de establecimiento la cual fue superada con un alto
porcentaje de sobrevivencia siguiendo el protocolo de esterilización con etanol
al 96% e hipoclorito de sodio al 2,5% las cuales fueron cultivadas en medio
Murashige & Skoog. En la fase de multiplicación se usaron 3 concentraciones
de ANA y 5 concentraciones de BAP, siendo el de mejor respuesta la combinación
de 0,25 µM ANA + 2 µM BAP con 15.8 brotes, los mismos que alcanzaron 9 mm de
tamaño, 2,7 hojas en promedio por explante y no presentaron formación de callo,
posteriormente se realizó dos subcultivos de este tratamiento cuya tasa de multiplicación
resultó 1:17 brotes en 30 días.
Palabras clave: Polylepis
rugulosa;
propagación in vitro; reguladores de crecimiento; establecimiento;
multiplicación.
ABSTRACT
Within the framework of the
forestry research and afforestation program of Sociedad Minera Cerro Verde,
this study was carried out, in order to establish a protocol for vegetative
propagation using the in vitro propagation technique of queñoa buds (Polylepis
rugulosa Bitter) for their conservation, as it is an endangered forest
species, for which the cultivation was carried out with different
concentrations of auxin and cytokinin in the in vitro multiplication phase,
passing through the establishment phase which was overcome with a high
percentage of survival following the protocol sterilization with 96% ethanol
and 2.5% sodium hypochlorite which were cultured in Murashige & Skoog
medium. In the multiplication phase, 3 concentrations of ANA and 5 concentrations
of BAP were used, with the best response being the combination of 0.25 µM ANA +
2 µM BAP with 15.8 shoots, the same ones that reached 9 mm in size, 2.7 leaves
on average per explant and did not show callus formation, subsequently two
subcultures of this treatment were carried out, whose multiplication rate was
1:17 shoots in 30 days.
Keywords: Polylepis
rugulosa;
in vitro propagation; growth regulators; establishment; multiplicaction.
Recibido: 04-05-2021.
Aceptado: 07-06-2021.
El género Polylepis se encuentra con la mayor
distribución de forma natural en Perú (Mamani, 2018), se tiene un reporte de 19
especies, considerándose, así como el país que presenta la más alta diversidad
de este género, de la cual resulta Cusco y Ayacucho con la mayor cantidad de
especies presentes en dichas zonas (Mendoza & Cano, 2011).
Los árboles de Polylepis tienen una corteza y
hojas que les permite adaptarse a extremas condiciones climáticas de la región
de los Andes (Zapata et al., 2012). Son importantes dado que albergan
diferentes especies de flora y fauna (Ames et al., 2019), en especial aves
(Ríos et al., 2018), así mismo la presencia de este género reduce los efectos
del cambio climático y regula los recursos hídricos (Mamani, 2018). Sin embargo
los peligros que afronta este género está relacionado con las actividades
humanas y las perturbaciones (Ríos et al., 2018), haciéndolos cada vez más
vulnerables debido a la deforestación y el pastoreo (Zutta et al., 2012). Por
otro lado, las semillas que pudieron colectar Canales & Huarasa (2020) de
la especie Polylepis incana fueron sometidas a tres tratamientos,
obteniendo un 9% de poder germinativo del tratamiento con agua residual. De
igual manera Vega et al. (2018), encontraron porcentajes de germinación entre
2% y 19% en tres especies de Polylepis. Lo cual demuestra que la queñoa es una
especie que presenta dificultad en su propagación por semillas botánicas.
De acuerdo a la información
obtenida del cultivo in vitro del género Polylepis se encontró
trabajos realizados por Mamani et al. (2015) y Taborga et al. (2016) que
desarrollaron la propagación hasta la fase de multiplicación en las especies Polylepis
pepei y Polylepis besseri respectivamente, mientras que Vega et al.
(2007) desarrollaron hasta la fase de enraizamiento en la especie Polylepis
tomentella, de todo estos trabajos realizados por diferentes autores se
pudo revisar que obtuvieron entre 3,1 y 5 brotes por explante en la fase de
multiplicación.
Mediante la técnica de
propagación in vitro, es posible desarrollar un protocolo de propagación que
permita su conservación y producción masiva de plantas en el menor tiempo
posible y en cualquier época del año lo cual favorecerá la reforestación y
restauración de bosques de queñoa.
Por todo lo expuesto el
objetivo de esta investigación fue generar un mayor número de brotes mediante
la combinación adecuada de la auxina (ANA) y citoquina (BAP).
El presente trabajo se desarrolló en el
Laboratorio de Biotecnología de Sociedad Minera Cerro Verde, distrito de
Uchumayo, Arequipa, Perú; llevándose a cabo en tres etapas:
Etapa 1
Extracción de los explantes
El material vegetal que se utilizó provino de
plantas madre de 90 cm de altura promedio, aclimatadas bajo condiciones de
vivero (Figura 3a), donde se les realizó tratamientos previos como
lavado de hojas, aplicación de fungicida y bioestimulantes.
Etapa 2 Establecimiento
Desinfección de explantes
Se extrajo brotes idealmente juveniles entre 4 y 5
cm de longitud promedio y que no presenten síntomas de enfermedades, los cuales
fueron colectados el mismo día que se realizó la siembra. Los explantes (Figura
3b) fueron lavados con abundante agua y jabón líquido por 3 minutos,
seguidamente inmersos en etanol al 90% durante 1 minuto en constante agitación,
luego sumergidos en solución de hipoclorito de sodio al 2,5% por 5 minutos y
finalmente se enjuagó tres veces con agua destilada esterilizada.
Siembra del material vegetativo
Se sembraron 5 yemas por frasco (Figura 3d),
se empleó el medio Murashige & Skoog al cual se agregó 3% de sacarosa y
0,6% de agar, el pH fue ajustado a 5,7; una vez concluida la siembra, los recipientes
de plástico fueron colocados durante 4 días a oscuridad absoluta y de esta
manera reducir el nivel de fenolización, para su evaluación se utilizó la
escala cualitativa que empleó Concepción et al. (2005) adaptado de Ramírez y
Salazar (1998), transcurrido el tiempo estos fueron colocados en la cámara de
incubación a una temperatura de 25°C con un fotoperiodo de 16 horas de luz y 8
horas de oscuridad. Dos semanas después de la siembra se efectuó la remoción de
sustancias fenólicas de la base de los explantes, en esta fase se evaluó en
porcentaje de contaminación y fenolización.
Etapa 3 Multiplicación
Se emplearon explantes provenientes de la fase de
establecimiento los cuales fueron cultivados en el medio de cultivo nutritivo
de Murashige & Skoog al cual se le adicionó diferentes concentraciones de
ANA (0, 0,25 y 0,5µM) y BAP (0, 2, 4, 6 y 8µM) bajo un Diseño de Bloques
Completamente al Azar con arreglo factorial 5 x 3, distribuidos en 15
tratamientos, 5 repeticiones y 75 unidades experimentales, con análisis de
varianza (ANOVA) y se estableció diferencias significativas con la prueba de
especificidad de LSD Fisher para los caracteres cuantitativos. Para obtener los
datos del peso de masa de los explantes/frasco (Tabla 3) se realizó 4
evaluaciones a partir de la semana 3 con el uso de una balanza analítica y la
evaluación final con respecto a las variables: número de brotes, longitud de
brotes, número de hojas y formación de callo se realizó a los 45 días después
de la siembra.
Fase
de establecimiento
Se pudo apreciar que los
explantes presentaron únicamente contaminación del tipo fungoso en un 9.8%
(bajo), debido a que
el material previamente fue instalado bajo condiciones de vivero. Cabe
mencionar que para reducir el porcentaje de contaminación se debe realizar
tratamientos previos a las plantas madre de las que se extraerán los explantes,
como lo concluyen Jones y Flores-Mora (2007): un buen manejo agronómico del
material en invernadero disminuye la presencia de microorganismos, como también
lo indicaron Cancino-Escalante et al. (2015); Abdelnour-Esquivel y Muñoz (2005)
mencionan que los explantes provenientes de estacas mantenidas en vivero
hicieron que los desinfectantes como el hipoclorito de calcio e hipoclorito de
sodio sean efectivos.
Sigarroa-Rieche y
García (2011), Martínez-Cruz et al. (2013) y Núñez et al. (2017) indican que
antes del proceso de desinfección con etanol e hipoclorito de sodio, lavaron
sus explantes con agua y jabón líquido comercial. Roca y Mroginski (1991)
sugieren que lavar los explantes con agua y detergente mejora la desinfestación
de explantes; del mismo modo se realizó en el presente trabajo.
Se utilizó el hipoclorito de sodio (NaClO) como
desinfectante ya que tiene la ventaja de enjuagarse más fácilmente de las
superficies después de la esterilización y así eliminar los agentes
contaminantes no deseados. La solución de hipoclorito de sodio (NaClO) a
concentraciones entre 1% a 3 % son las más usadas en el proceso de
esterilización de explantes siempre que no afecte el estado de los tejidos
(Borges et al., 2009).
Fenolización / Oxidación
Se pudo evitar que la
fenolización sea muy alta al someter los explantes sembrados a oscuridad
durante 4 días, ya que la queñoa, por ser leñosa, tiene la tendencia a
fenolizar, se obtuvo 89.16% de fenolización, con un nivel de poco fenolizado. Méndez-Álvarez
et al. (2014) realizaron un trabajo en donde después de haber desinfectado y
sembrado los explantes, los colocaron en un cuarto de crecimiento donde
permanecieron en oscuridad durante una semana, posteriormente los transfirieron
a una intensidad lumínica de 2000 lux con un fotoperiodo de 16 horas luz.
En el trabajo
desarrollado por Jacinto (2018), menciona que todos sus tratamientos
presentaron oxidación, atribuyendo al daño que sufren los tejidos en el momento
de realizar los cortes de la parte basal del material lo cual concuerda con el
presente trabajo, ya que se obtuvo también un porcentaje alto de fenolización
en la parte basal de los explantes (Figura 3c).
Azofeifa (2009)
alude que existen varias estrategias
para evitar la oxidación de los explantes. En primer lugar está la prevención y
disminución de las causas que provocan o estimulan el estrés oxidativo. Sin
embargo si no se pudiera evitar se puede recurrir a otros procedimientos con la
finalidad de controlar el proceso oxidativo; por lo cual, transcurridas dos semanas después de la
siembra, se procedió con la remoción de sustancias fenólicas, se pudo notar
que, al realizar esta actividad, los explantes empezaban a desarrollarse ya que
el estar en el medio inicial no les permitía desarrollarse.
Fase de
multiplicación
Crecimiento en peso
En la Tabla 1 se
muestra los datos obtenidos del peso de masa de brotes/explante de las semanas
3, 4, 5 y 6 en la fase de multiplicación, mediante la prueba estadística ANOVA
(p > 0,05)
nos indica que existe diferencia significativa entre los tratamientos en las
semanas 3, 4, 5 y 6; en tanto la prueba de especificidad de LSD Fisher muestra
que durante la semana 3 el tratamiento 0 µM ANA + 2 µM BAP
presentó el más alto valor, con 0.83 g en comparación al restos de
tratamientos; durante la semana 4 los tratamientos 0 µM ANA + 2 µM BAP y 0,25
µM ANA + 2 µM BAP presentaron los más altos valores de peso promedio con 1,08 y
1,24 g respectivamente; en la semana 5 el tratamiento 0,25 µM ANA + 2 µM BAP
presentó el más alto valor en peso con 2,25 g y finalmente en la semana 6, el
tratamiento 0,25 µM ANA + 2 µM BAP presentó el más alto valor en peso con 4,22
g con respecto al resto de tratamientos. Los pesos iniciales a los 21 días podrían
estar influenciados por el tamaño de los explantes; sin embargo, al momento de
la siembra se trató de emplear yemas de un tamaño uniforme.
Tabla 1
Peso de masa de
brotes/explante de Polylepis rugulosa Bitter en 15 tratamientos
combinados de ANA y BAP
|
Tratamiento
|
ANA (µM)
|
BAP (µM)
|
Peso promedio (g) en semana
|
|
3
|
4
|
5
|
6
|
|
T1
|
0 +
|
0
|
0,67 bc
|
0,83 b
|
1,04 def
|
1,28 fg
|
|
T2
|
|
2
|
0,83 a
|
1,08 a
|
1,77 b
|
2,88 b
|
|
T3
|
|
4
|
0,48 ef
|
0,64 cd
|
1,14 cde
|
1,64 cdef
|
|
T4
|
|
6
|
0,41efgh
|
0,66 cd
|
1,05 ef
|
1,72 cd
|
|
T5
|
|
8
|
0,36 fgh
|
0,54 def
|
0,85 fg
|
1,35 efg
|
|
T6
|
0,25 +
|
0
|
0,34gh
|
0,36 g
|
0,37 h
|
0,39 h
|
|
T7
|
|
2
|
0,77ab
|
1,24 a
|
2,25 a
|
4,22 a
|
|
T8
|
|
4
|
0,60 cd
|
0,86 b
|
1,33 c
|
1,94 c
|
|
T9
|
|
6
|
0,29 h
|
0,46 efg
|
0,67 g
|
1,17 g
|
|
T10
|
|
8
|
0,53 df
|
0,84 b
|
1,27 cd
|
1,76 cd
|
|
T11
|
0,5 +
|
0
|
0,36 fgh
|
0,38 fg
|
0,39 h
|
0,44 h
|
|
T12
|
|
2
|
0,37 fgh
|
0,55 de
|
0,88 f
|
1,49 defg
|
|
T13
|
|
4
|
0,46 efg
|
0,65 cd
|
0,95 ef
|
1,34 efg
|
|
T14
|
|
6
|
0,37 fgh
|
0,59 cde
|
0,97 ef
|
1,69 cde
|
|
T15
|
|
8
|
0,50 de
|
0,72 bc
|
1,10 de
|
1,94 c
|
a, b, c… corresponden a la prueba
LSD Fisher α=0,05.
Se puede observar
(Tabla 1) que cada semana que transcurría se producía un incremento del peso
notándose que los tratamientos 0 µM ANA + 2 µM BAP y 0,25 µM ANA + 2 µM BAP
mantuvieron los más altos valores de peso durante las semanas evaluadas por
otro lado los tratamientos 0,25 µM ANA + 0 µM BAP y 0,5 µM ANA + 0 µM BAP
presentaron los valores más bajos durante las semanas evaluadas y no existe
mucha diferencia en el aumento de peso durante las 4 semanas evaluadas, a
diferencia del resto de tratamientos, lo cual demuestra que al no aplicar
citoquinina BAP a estos tratamientos no hubo división celular por lo tanto
tampoco formación de nuevos brotes lo que hizo que el aumento en peso no sea
tan notorio en cada semana y al contener únicamente auxina ANA se mantuvo la
dominancia apical.
Posteriormente se
realizó el análisis de correlación entre el peso y el número de brotes
obteniéndose un coeficiente de correlación de 0.92 entre las variables lo cual
indica que es una relación lineal positiva y fuerte según las consideraciones
de interpretación de Martínez et al. (2009). Dicho resultado se corrobora con
el trabajo realizado por Colmenares y Giménez (2003), en el cual para el banano
cv. ‘Williams’ obtuvieron la mayor cantidad de brotes (8,4) y mayor peso fresco
por explante en SIT (19,9 g) que en medio de cultivo sólido que proporcionó la
menor cantidad de brotes (2,4) y peso fresco (4,4 g) ambos datos lo obtuvieron
a los 30 días de su cultivo, mencionando que existe una correlación directa
entre el peso fresco y el incremento del número de brotes. En la figura 1
se puede apreciar que a medida que incrementa el peso también se incrementa el
número de brotes.

Figura 1. Relación
lineal positiva y fuerte entre las variables de peso y número de brotes.
Número de brotes
El número de brotes
obtenidos de los quince tratamientos durante el primer subcultivo (Figura 3f)
en la fase de multiplicación mediante la prueba estadística ANOVA (P > 0,05), nos indica que
existe diferencia significativa entre los tratamientos, la prueba de
especificidad de LSD Fisher nos indica que el tratamiento 0,25
µM ANA + 2 µM
BAP
muestra los
mejores resultados con 15,8 brotes (Figura 2), lo cual concuerda con lo que
menciona Pierik (1997): las citoquininas generalmente promueven la división
celular, especialmente si se agrega con una auxina; también coincide con los
trabajos que desarrollaron Mamani y Murillo (2020), Medina et al. (2014) y
Castellanos et al. (2006) obteniendo en la interacción de citoquinina y auxina
que los explantes empleados promovieron mayor número de brotes, siendo
necesaria la presencia de ambos reguladores de crecimiento para la fase de
multiplicación.
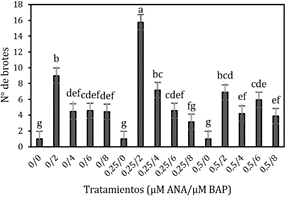
Figura 2. Número de brotes a los 45 días en la fase de
multiplicación in vitro de yemas de Polylepis rugulosa (queñoa)
en diferentes concentraciones de citoquina y auxina.
Es necesario mencionar que todos los tratamientos
que contenían citoquinina BAP presentaron formación de nuevos brotes (Figura
3e) a pesar de que algunos no contenían auxina, lo cual se relaciona con los
resultados mostrados por Torres y Suárez (2013), en los que, para la aplicación
de citoquininas en su caso de estudio emplearon el BAP, vieron que es necesaria
para incrementar el número de brotes en la fase de multiplicación a partir de
explantes en condiciones in vitro. Por lo tanto, los tratamientos que tienen
cero concentraciones de citoquinina presentaron nula formación de nuevos brotes
comprobando que las citoquininas promueven la formación de brotes axilares
(Pierik, 1997). Así mismo Solis et al. (2011) afirma que el número de brotes es
una variable importante en la fase de multiplicación ya que permitirá obtener
la tasa de multiplicación.
Con relación a los resultados según factores
principales, al realizar la prueba de especificidad de LSD Fisher se observa
(Tabla 2) que la mejor respuesta para el factor principal citoquinina (BAP) es de
10,55 brotes por explante de la concentración 2 µM diferente estadísticamente
al resto de las concentraciones, mientras que el menor valor lo representa la
concentración 0 µM de BAP, con un promedio de 1 brote por explante similar
resultado obtuvieron Rodríguez et al. (2003) en la propagación in vitro de
Artemisia absinthium en donde el tratamiento sin BAP (testigo), obtuvo
el menor e igual número de brotes en cada uno de los períodos evaluados (20, 30
y 40 días). Respecto a los resultados para el factor principal auxina (ANA) la
concentración que permitió la multiplicación de brotes es de 0,25 µM con 6,33
brotes por explante, mientras que las concentraciones 0 µM y 0,50 µM
presentaron 4,70 y 4,38 brotes / explante respectivamente, y ambas
concentraciones no presentan diferencia significativa.
Tabla 2
Número de brotes a los 45 días después de la
siembra a nivel de factores principales en la fase de multiplicación in vitro
de yemas de Polylepis rugulosa Bitter (queñoa)
|
Factores
principales
|
|
Citoquina
|
Concentración
BAP
|
Promedio
|
Auxina
|
Concentración
ANA
|
Promedio
|
|
C2
|
2 µM BAP
|
10,55 a
|
A2
|
0,25 µM ANA
|
6,33 a
|
|
C3
|
4 µM BAP
|
5,28 b
|
A1
|
0,00 µM ANA
|
4,70 b
|
|
C4
|
6 µM BAP
|
5,01 b
|
A3
|
0,50 µM ANA
|
4,38 b
|
|
C5
|
8 µM BAP
|
3,83 b
|
|
|
|
|
C1
|
0 µM BAP
|
1,00 c
|
|
|
|
a,b,c…
corresponden a la prueba LSD Fisher α=0,05.
En el presente trabajo también se determinó la
tasa de multiplicación para ello se realizó dos siembras adicionales del
tratamiento 0,25µM ANA+ 2µM BAP ya que presentó la mayor cantidad de brotes, en el segundo
subcultivo se obtuvo un promedio de 15,4 yemas por cada yema sembrada
inicialmente, mientras que el tercer subcultivo 19,5 yemas por cada yema
sembrada; finalmente con los tres subcultivos realizados se sacó el promedio
obteniendo una tasa de multiplicación de 1:17. El número de brotes es una
variable importante en la fase de multiplicación ya que a mayor número de
brotes se tendrá una mayor tasa de multiplicación

Figura 3. Secuencia de la micropropagación en Polylepis
rugulosa Bitter. (a) Plantas madre, (b) Brotes diseccionados,
(c) Poca fenolización y traslado de yemas (d) Explantes en medio
de multiplicación, (e) Proliferación de brotes/explante/frasco, (f)
Brotes aislados por explante.
Longitud
de brotes
Mediante la prueba estadística ANOVA (P > 0,05)
muestra que existe diferencia significativa con respecto a la altura de los
brotes, la prueba de especificidad de LSD Fisher indicó que los trata-mientos 0
µM ANA + 0µ
M BAP
y 0,5
µM ANA + 0 µM BAP presentaron los
mejores resultados (31 y 27 mm respectivamente), seguido del tratamiento 0,25
µM ANA + 0 µM BAP con 20 mm; mientras
que el resto de los tratamientos no presentaron diferencias significativas
entre sí (Figura
4).
Se pudo observar que aquellos tratamientos en los
cuales estuvo presente la citoquinina BAP presentan alturas entre 8,6 mm y 12
mm como máximo, mientras que aquellos tratamientos en los cuales no se aplicó
citoquinina BAP presentan los valores más altos en altura, lo cual demuestra
que sin la incorporación de citoquinina obtenemos una mayor altura foliar
mientras que la presencia de citoquinina en los tratamientos la altura
disminuye, coincidiendo con lo que menciona Pierik (1997) sobre cómo las
citoquininas disminuyen la dominancia apical, y también con Arredondo (2000),
citado por Uribe y Cifuentes (2004), señalando que para obtener explantes de
mayor longitud, es necesario que el medio de cultivo sea suplementado con
una menor concentración de citoquininas, ya que con esto se logra disminuir la
división celular y favorecer la elongación del tejido por la acción de las
auxinas, los resultados obtenidos concuerdan con lo que obtuvo Cob et al.
(2016), que al bajar la concentración de citoquinina en la relación AIB/BAP, el
tamaño de los explantes de Quillaja Saponaria aumentaron; la altura
optima lo alcanzaron con el tratamiento T9 (1,5/0,5 mg L-1 de
AIB/BAP). En contraste, los microtallos que fueron cultivados en el tratamiento
T5 (1,0/3,0 mg L-1 de AIB/BAP) no elongaron, y observaron
explantes en forma de roseta.

Figura 4. Longitud de brotes a los 45 días en la fase de
multiplicación in vitro de yemas de Polylepis rugulosa (queñoa)
en diferentes concentraciones de citoquinina y auxina.
en donde encontraron
que el tamaño de los explantes aumentaron al bajar la concentración de
citoquinina en la relación AIB/BAP
El tratamiento 0 µM ANA+ 0 µM
BAP presentó la mayor altura (31 mm) en comparación al resto
tratamientos, resulta importante mencionar que este tratamiento no contiene
reguladores de crecimiento lo cual coincide con los resultados obtenidos por Rodríguez et al. (2003), donde evaluaron
el comportamiento de la altura de los brotes, en el cual el tratamiento testigo
(T-1), sin reguladores, obtuvo los mayores valores en 3 períodos evaluados (20,
30 y 40 días) que fueron estadísticamente diferentes entre ellos y con el resto
de los tratamientos y similar resultado se encontró con el trabajo
realizado por Gutierrez-Rozeti y Gonzales (2019).
Se pudo notar que los tratamientos que obtuvieron
los más altos valores en cuanto a la variable longitud de brote presentaron los
valores más bajos respecto a la variable número de brotes, semejante resultado
fue obtenido por Vega et al. (2007), ya que observaron que el
grupo 1A presentó el mayor promedio en cuanto a la altura de explantes y al
mismo tiempo el menor número de brotes a los 30 días de evaluación. Esta
observación demuestra una relación inversamente proporcional respecto a estas
variables; es decir, que a mayor longitud del explante se registró una menor
formación de brotes y viceversa; no obstante, estos resultados difieren de los
encontrados por Mamani et al. (2015) debido a que el tratamiento que presentó
el mayor número de brotes mostró a su vez una mayor longitud y número de hojas.
Número de hojas
Con respecto al número de hojas la prueba
estadística ANOVA (P > 0,05) nos indica que existe diferencia significativa
entre los tratamientos, la prueba de especificidad de LSD Fisher muestra que
los tratamientos 0 µM ANA + 0 µM BAP; 0,25
µM ANA + 0 µM
BAP y 0,5
µM ANA + 0 µM BAP presentaron los más
altos valores, con 4,7; 4,6 y 4,5 hojas en promedio por brote respectivamente (Figura 5).
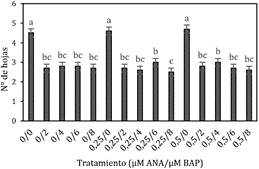
Figura 5. Número de hojas a los 45 días en la fase de
multiplicación in vitro de yemas de Polylepis rugulosa (queñoa)
en diferentes concentraciones de citoquinina y auxina.
Los tratamientos que obtuvieron los más altos
valores en cuanto a la variable longitud de brote también presentaron los más
altos valores respecto a la variable número de hojas, lo cual permite afirmar
para el caso del estudio realizado que el número de hojas es directamente
proporcional a la longitud de brotes, lo que quiere decir que a mayor longitud
mayor número de hojas, similar resultado encontrado por Villa y Arbeláez (2019) con relación a las variables de altura y número
de hojas en uno de sus tratamientos empleados. Respecto a las variables número
de brotes y número de hojas se observaron resultados inversamente
proporcionales, lo cual difiere con lo obtenido por Vega et al. (2007) en donde
observaron una relación directamente proporcional entre la
formación de brotes y el número de hojas diferenciadas en los explantes
evaluados.
Formación
de callo
En la Figura 6 se muestra los porcentajes de la
formación de callo que presentaron los explantes sometidos a diferentes
concentraciones de reguladores de crecimiento (auxina y citoquinina), se puede
notar que la mayoría de tratamientos no presentaron formación de callo a
excepción de los tratamientos 0,25 µM ANA + 6 µM BAP; 0,25 µM ANA + 8 µM BAP;
0,5 µM ANA + 0 µM BAP y 0,5 µM ANA + 4 µM BAP con una ligera formación de
callo, sin embargo al realizar el análisis estadístico la prueba estadística
ANOVA (P > 0,05) indica que todos los tratamientos son iguales, por lo que
cada uno de los tratamientos pueden ser empleados.
Roca y Mroginski (1991), mencionan que la
propagación clonal trae consigo producir un nuevo individuo con los mismos
genes de su progenitor y hasta el momento los explantes más usados en la propagación
in vitro son puntas de tallos y yemas laterales. Por tal motivo en la
micropropragación muy pocas veces se usan callos. Cabe mencionar que para el
caso del estudio realizado está orientado a producir nuevas plantas a través de
la organogénesis directa es decir producir una nueva planta a partir de yemas
sin formación de callo.
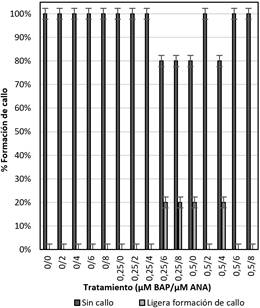
Figura 6. Porcentaje de formación de callo a los 45 días en
la fase de multiplicación in vitro de yemas de Polylepis rugulosa
(queñoa) en diferentes concentraciones de citoquinina y auxina.
La concentración de citoquinina de 2 µM BAP en
combinación con la auxina de 0,25 µM ANA generó la mayor proliferación de
brotes de 1:17. Se recomienda realizar el subcultivo de explantes a los 60 días
en la fase de multiplicación para probar superar el número de brotes obtenidos.
Realizar investigaciones en otras especies de queñoa, con la concentración que
permitió obtener la mayor cantidad de brotes en Polylepis rugulosa Bitter,
para probar el mismo resultado en la fase de multiplicación.
Se agradece el financiamiento de esta
investigación a Sociedad Minera Cerro Verde, que permitió se haga posible el
desarrollo del trabajo de investigación bajo el contrato N° 37031903237 – CLS-236-2019.
También se agradece a Eduardo Molinari-Novoa (Chess Consulting & Project)
por sus comentarios al presente manuscrito.
REFERENCIAS
BIBLIOGRÁFICAS
Abdelnour-Esquivel, A., & Muñoz, A. (2005).
Micropropagación de teca (Tectona grandis Lf). Kurú: Revista Forestal
(Costa Rica) 2(5), 2005.
Ames-Martínez, F. N., Quispe Melgar, H. R., Zúñiga López, D.
G., Segovia Salcedo, M. C., & Kessler, M. (2019). Bosques de
Polylepis: Biodiversidad en la región central del Perú. Universidad
Continental.
Azofeifa, Á. (2009). Problemas de oxidación y oscurecimiento
de explantes cultivados in vitro. Agronomía mesoamericana, 20(1),
153-175.
Borges, G. M., Estrada, A. E., Pérez, R. I., Meneses, R. S.
(2009). Uso de distintos tratamientos de desinfección en el cultivo in vitro
de Discorea alata L. clon caraqueño. Cuba.
Canales, Á., & Huarasa, Y. R. (2020). Poder germinativo
de Polylepis incana con aplicación de diferentes tratamientos de
agua. Revista Cubana de Ciencias Forestales, 8(3),
495-506.
Cancino-Escalante, G. O., García, E. Q., Villamizar, C. E.,
& Carvajal, C. D. (2015). Propagación in vitro de materiales seleccionados
de Rubus glaucus Benth. (mora de Castilla) en la provincia de Pamplona,
región nororiental de Colombia. Revista Colombiana de Biotecnología, 17(2),
7-15.
Castellanos, O., Rodríguez, A., Rodríguez, J., y Rodríguez,
B. (2006). Organogénesis indirecta y enraizamiento in vitro de Paulownia
elongata. e-Gnosis, 4.
Cob, J., Ríos, D., Sabja, A., Cartes, P., & Sánchez, M.
(2016). Organogénesis directa para la propagación in vitro de Quillaja
saponaria Molina en Sudamérica Austral. Revista mexicana de
ciencias forestales, 7(34), 57-68.
Colmenares, M., & Giménez, C. (2003). Multiplicación in
vitro de Musa spp. mediante sistema de inmersión temporal. Revista
de la Facultad de Agronomía, 20(4), 468-477.
Concepción, O., Nápoles, L., Pérez, A. T., Peralta, N.,
Hernández, M., & Trujillo, R. (2005). Efecto de tres antioxidantes en el cultivo
in vitro de ápices de guayaba (Psidium guajava L.). Relación entre el
origen del explante y el contenido de compuestos fenólicos. Cultivos
tropicales, 26(1), 33-39.
Gutierrez-Rosati, A.; Gonzales, P. (2019). Reguladores de
crecimiento en el cultivo in vitro de tres cultivares portainjertos de
vid (Vitis vinifera L.) para su uso en la industria del pisco. Scientia
Agropecuaria, 10(4), 461-468.
Jacinto, M. (2018). Evaluación de tres niveles de auxinas y
citoquininas para la obtención de plantas madre de rosa (Rosa sp.)
variedad ‘Freedom’ en condiciones in vitro. Apthapi, 4(2),
1073-1081.
Jones, F.,
& Flores-Mora, D. M. (2007). Establecimiento in vitro y pruebas
preliminares de micropropagación en medio semisólido y líquido de frambuesa (Rubus
idaeus L.). Revista Tecnología En Marcha, 20(3),
46.
Mamani, M. S. (2018). Diversidad genética de poblaciones de Polylepis
canoi W. Mendoza y Polylepis rodolfo-vasquezii L. Valenzuela &
L. Villalba mediante el uso de microsatélites (SSR) en la región Junín.
Mamani, B., Rocabado Koya, P., & Rey Ortíz, L.
(2015). Establecimiento y multiplicación in vitro de Polylepis pepei
(No. CIDAB-S531-I51r-5/2015). Instituto Nacional de Innovación Agropecuaria y
Forestal (Bolivia).
Mamani, B., & Murillo, R. A. (2020). Micropropa-gación de
dos variedades de frutilla (Fragaria Ananassa Duch.) En diferentes medios de
cultivo. Revista de Investigación e Innovación Agropecuaria y de
Recursos Naturales, 7(1), 69-78.
Martínez, R. M., Tuya, L. C., Martínez, M., Pérez, A., &
Cánovas, A. M. (2009). El coeficiente de correlación de los rangos de Spearman
caracterización. Revista Habanera de Ciencias Médicas, 8(2), 0-0.
Martínez-Cruz,
N. S., Arévalo-Niño, K., Verde-Star, M. J., Oranday-Cárdenas, A.,
Rivas-Morales, C., Treviño-Neávez, J., & Morales-Rubio, M. E. (2013).
Germinación in vitro e inducción de callo en Rubus adenotrichus Schltdl.
Polibotánica, (35), 99-107.
Medina, M., Mosquera, H., & Aguilar, C. (2014).
Micropropagación clonal y enraizamiento ex vitro de tres cultivares de piña Ananas
comosus (L. Merr.) del Chocó, Colombia. Revista Biodiversidad
Neotropical, 4(2), 133-140.
Méndez-Álvarez, D., & Abdelnour-Esquivel, A. (2014).
Establecimiento in vitro de Terminalia amazonia (Gmel.) Excell. Revista
Forestal Mesoamericana Kurú, 11(27), 7-21.
Mendoza, W., & Cano, A. (2011). Diversidad del género Polylepis
(Rosaceae, Sanguisorbeae) en los Andes peruanos. Revista peruana de
biología, 18(2), 197-200.
Núñez, J. E. N., Quiala, E., de Feria, M., Mestanza, S.,
& Teanga, S. (2017). Propagación in vitro de Caesalpinia spinosa
(Mol.) O. Kuntz a partir de yemas axilares de árboles plus seleccionados. Biotecnología Vegetal, 17(2).
Pierik, R. (1997). In vitro culture of
higher plants. Springer science & business media.
Ríos, C. S. S., Rodewald, A. D., & Morales,
L. V. (2018). Ecología y conservación de las aves asociadas con
Polylepis: ¿qué sabemos de esta comunidad cada vez más vulnerable? Ecología
Austral, 28(1-bis), 216-228.
Roca, W. M., & Mroginski, L. (1991). Cultivo de tejidos
en la Agricultura: Fundamentos y aplicaciones. (CIAT) No. 151.
Rodríguez, H., Hechevarría, I., Rodrígue, C. A., &
Rivera, M. M. (2003). Propagación in vitro de Artemisia absinthium L. en
Cuba. Revista Cubana de Plantas Medicinales, 8(1), 0-0.
Sigarroa-Rieche, A. K., & García-Delgado, C. L. (2011).
Establecimiento y multiplicación in vitro de mora de castilla (Rubus glaucus
Benth.) variedad sin espinas, mediante ápices meristemáticos. Acta
agronómica, 60(4), 347-354.
Solis, R., Olivera, J., & La Rosa, R. S. (2011).
Propagación in vitro de Carica papaya var. PTM-331 a partir de
meristemos apicales. Revista peruana de biología, 18(3),
343-348.
Taborga, A. D. A., León, R. M., Poma, D. V., García, V.,
& Rojas, J. J. V. (2016). Desarrollo de un protocolo para el
establecimiento in vitro de Kewiña (Polylepis besseri, Hieron.) a partir
de yemas apicales. Revista Científica de Investigación Info-Iniaf, 1(8),
43-49.
Torres, L. A., & Suarez, I. E. (2013). Propagación in
vitro de Acacia mangium Willd. Biotecnología en el Sector
Agropecuario y Agroindustrial: BSAA, 11(1), 81-87.
Uribe, M. & Cifuentes, L. (2004). Aplicación de técnicas
de cultivo in vitro en la propagación de Legrandia concinna. Bosque
(Valdivia), 25(1), 129-135.
Vega, C. K., Villegas, C. G., Rocabado, P. A., Quezada, J.
A., López, M. Y., & Quevedo, A. W. (2018). Biología reproductiva de tres
especies de Polylepis (P. neglecta, P. incarum y P.
pacensis), con énfasis en su comportamiento germinativo. Ecología
Austral, 28(1-bis), 310-324.
Vega, C., Bermejo-Franco, J. C., & Villegas-Alvarado, G.
(2007). Propagación masiva de Polylepis tomentella Weddell ssp. nana
mediante técnicas de cultivo in vitro. Ecología en Bolivia, 42(2),
102-120.
Villa, R., & Arbeláez, L. M. (2019). Micropropagación in
vitro de rosa rosa sp. a partir de yemas axilares y respuesta
callogénica. Revista de la Asociación Colombiana de Ciencias Biologicas, 1(31),
10-17.
Zapata, F., Dourojeanni, P., Gagliardi, G., & Cordero, D.
(2012). El Clima Cambia, Cambia Tú También. Adaptación al cambio climático en
comunidades locales del Perú. Lima, Sociedad Peruana de Derecho
Ambiental.
Zutta, B. R., Rundel, P. W., Saatchi, S., Casana, J. D.,
Gauthier, P., Soto, A. & Buermann, W. (2012). Prediciendo la distribución
de Polylepis: bosques Andinos vulnerables y cada vez más
importantes. Revista Peruana de Biología, 19(2),
205-212.