Aislamiento
e identificación de Leishmania sp de Amblyomma naponense y Amblyomma
humerale colectadas de Pecari tajacu, Tayassu pecari y Chelonoide
denticulata
Isolation
and identification of Leishmania sp of Amblyomma naponense and Amblyomma
humerale collected from Pecari tajacu, Tayassu pecari and Chelonoide
denticulata
Jesús Rojas-Jaimes 1,
2, *; Daniel Zarate-Rendon 3; Gino Montoya-Arauco 4
1
Facultad de Ciencias de la Salud, Universidad Privada del Norte, Lima, Perú.
2
Escuela de Medicina Humana, Universidad Científica del Sur, Lima, Perú.
3
Laboratorio de Parasitología, Facultad de Zootecnia, Universidad Nacional Agraria
La Molina, Lima, Perú.
4
Escuela de Medicina Veterinaria, Universidad Científica del Sur, Lima, Perú.
*Autor
corresponsal: jesus.rojas.jaimes@gmail.com (J. Rojas-Jaimes).
ID
ORCID de los autores
J.
Rojas-Jaimes:  https://orcid.org/0000-0002-6910-9341 D.
Zarate-Rendon: https://orcid.org/0000-0003-3532-8084
https://orcid.org/0000-0002-6910-9341 D.
Zarate-Rendon: https://orcid.org/0000-0003-3532-8084
G.
Montoya-Arauco: https://orcid.org/0000-0003-1986-4557
RESUMEN
El objetivo del presente estudio fue aislar e
identificar a Leishmania de Amblyomma naponense y Amblyomma
humerale colectadas de Pecari tajacu, Tayassu pecari y Chelonoide
denticulata. Se colectaron garrapatas Amblyomma naponense y Amblyomma
humerale de Pecari tajacu, Tayassu pecari y Chelonoide
denticulata en Madre de Dios, Perú. Se extrajeron las vísceras y glándulas
salivales de las garrapatas y se hicieron observaciones directas al
microscopio, se realizaron cultivos en agar sangre bifásico y se inocularon en
almohadillas plantares e intraperitoneal de hámsteres. En ninguna de las
observaciones, cultivos y respuesta a las inoculaciones se aisló ni identificó
al parásito. Estos resultados muestran una investigación básica la cual sirve
para recomendar el uso de un número mayor de garrapatas colectadas de varios
animales como potenciales reservorios de Leishmania y el uso de técnica
de alta sensibilidad de detección del parásito como la reacción en cadena de
polimerasa (PCR).
Palabras clave: Leishmania; garrapata; aislamiento;
reservorio; vector.
ABSTRACT
The aim of this study was
to isolate and identify Leishmania from Amblyomma naponense and Amblyomma
humerale collected from Pecari tajacu, Tayassu pecari and Chelonoides
denticulata. Amblyomma naponense and Amblyomma humerale
ticks were collected from Pecari tajacu, Tayassu pecari and Chelonoides
denticulata in Madre de Dios, Peru. The viscera and salivary glands of the
ticks were extracted, and direct observations were made under the microscope.
Samples were cultured in biphasic blood agar and inoculated into the footpads
and intraperitoneal of hamsters. In none of the observations, cultures and responses
to inoculations was the parasite isolated or identified. These results show
basic research which serves to recommend the use of a larger number of ticks collected
from various animals as potential reservoirs of Leishmania and the use of a
highly sensitive parasite detection technique such as polymerase chain reaction
(PCR).
Keywords: Leishmania;
tick; isolation; reservoir; vector.
Recibido: 20-04-2022.
 Aceptado: 24-07-2022.
Aceptado: 24-07-2022.
2.1 Colecta de Garrapatas
Se coordinó con los cazadores de los
distritos de Planchon “Botijón”, Iberia e Iñapari (distrito de Bélgica) en
Madre de Dios, Perú, para la recolección de garrapatas en los animales que
hayan sido cazados. Los pobladores de las comunidades nativas realizan la
cacería de estos animales para autoconsumo.
Se colectaron 29 garrapatas de Pecari
tajacu en el distrito de Bélgica, 30 de Tayassu pecari y 10 de Chelonoide denticulata en el lapso de dos
semanas. Las garrapatas fueron transportadas vivas al laboratorio de
parasitología de la Universidad Agraria la Molina de la escuela de Zootecnia,
en frascos herméticos estériles, al laboratorio para la clasificación
taxonómica y la extracción de vísceras. La clasificación taxonómica se basó en
las claves de Barros-Battesti et al. (2006).
Por otro lado, y en forma casual, se
colectó una garrapata del brazo de uno de los cazadores del poblado de Botijón.
Se retiró todo resto de suciedad de las garrapatas con papel toalla y fueron
colocadas en placas Petri conteniendo 5 a 10 gotas de una solución de suero
fisiológico, antibiótico y antimicótico. Trabajando en el estereoscopio, se
diseccionaron las garrapatas con ayuda de microestiletes entomológicos para
exponer sus órganos internos. En el caso de las teleoginas más grandes fue
necesario realizar un piquete previo a la apertura le extremo posterior del
parásito.
2.2 Inoculación en hámsteres
Las garrapatas
se agruparon en pooles por especie y animal de colecta. Se extrajeron
las glándulas salivales y segmentos intestinales usando el estereoscopio Nikon
SMZ 800N, material que fue colocados en tubos Vacutainer®
conteniendo suero fisiológico más antibiótico y antimicótico. En un tubo de
ensayo de 4 ml se colocaron las vísceras y glándulas salivales de las 28
garrapatas colectados de Pecari tajacu, las cuáles una vez homogenizadas
se distribuyeron en tubos de 2 ml. Asimismo, se procedió en forma similar con
las vísceras y glándulas salivales de las 30 garrapatas colectados de Tayassu
pecari y las 10 garrapatas colectadas de Chelonoide denticulata.
Un tubo ensayo de cada par de pooles
se utilizó en la inoculación a hámsteres dorados hembras Mesocricetus
auratus (0,5 ml en cada pata). El contenido del segundo tubo de ensayo de
cada par fue usado para la inoculación intraperitoneal (0,5 ml), cultivo en
medio bifásico (1 ml) y microscopia (0,5 ml).
2.3 Observación microscópica y cultivo
Las muestras para microscopía se
colocaron en laminas portaobjetos y teñidas con Giemsa para la identificación
de tripanosomátidos usando un microscopio Nikon ECLIPSE Ei. Las láminas fueron
observadas a 400X y 1000X. Se realizaron cultivos de agar sangre bifásico
utilizando cloruro de sodio isotónico al 0,9% con penicilina 10 mil U/ estreptomicina
10 mg/ml y sangre desfibrinada de conejo. De cada muestra se hicieron tres
cultivos (repeticiones). Las muestras fueron incubadas a 25 °C por 4
semanas, realizándose observaciones dos veces por semana.
2.4 Observación histológica
Los
hámsteres tenían un mes de
edad y fueron acondicionados en jaulas individuales con ventilación natural y
cama de viruta en el bioterio de parasitología de la Facultad de Ciencias de la
Salud de la Escuela de Veterinaria de la Universidad Científica del Sur. En los hámsteres inoculados se evaluó el desarrollo de la enfermedad durante seis
meses, de lunes a viernes, excepto sábados y domingos. Al final del sexto
mes se tomaron muestras de biopsias a nivel hepático,
pulmón, riñón y bazo de 1 cm2, aproximadamente, para fijarlos en
formol y hacer láminas para los estudios histopatológicos a través de
coloración Hematoxilina/Eosina. En forma paralela, se utilizó un hámster como control positivo basado en la
inoculación de una solución de 2,34 x 106/ml de Leishmania (cepa
JM) en cultivo Schneider y como control negativo se utilizó un hámster
inoculado con una solución isotónica de cloruro de sodio al 0,9%. En ambos
casos, se inoculó 0,5 ml en
las almohadillas de las patas traseras y delanteras (2 ml total) y 0,5 ml vía
intraperitoneal.
Las garrapatas colectadas de T.
pecari (30 machos) y P. tajacu (28 machos) fueron clasificadas como Amblyomma
naponense y las colectadas de C. denticulata (10 machos) se
clasificaron como A. humerale.
En ninguna de
las muestras observadas mediante microscopio óptico del pool de glándulas
salivales y segmentos intestinales de las garrapatas se observaron
tripanosomátidos excepto en la garrapata colectada accidentalmente de una
persona en la que se encontraron tripanosomátidos en forma flagelada del pool
de glándulas salivales y segmentos intestinales (Figura 1).

Figura 1. Coloración
Giemsa de un extendido con vista directa al microscopio de la garrapata A.
naponense colectada de una persona en el poblado de Botijon/Las Piedras
visto a 400X donde se observa algunos flagelados.
Los cultivos fueron negativos para el
crecimiento de algún tripanosomátido flagelado, aunque se encontró un hongo Fusarium
y levaduras en la muestra de A. naponense colectado de Pecari tajacu (Figuras
2 y 3).
En los cortes
histopatológicos de las muestras provenientes de tejido hepático y bazo de los
hámsteres retados con la cepa JM de promastigotes de Leishmania sp
mostraron edema sinuoso de leve infiltrado inflamatorio linfocitario
distribuido en todo el parénquima y reacción inflamatoria, tipo pavimentoso
concordante con Leishmaniasis respectivamente. Las evaluaciones de las
demás muestras histológicas no fueron concluyentes.
Las garrapatas
colectadas de T. pecari y P. tajacu fueron clasificadas como A.
naponense y las colectadas de C. denticulata como A. humerale.
Estudios previos han demostrado la distribución especies del género Amblyomma
en animales silvestres como en Tapirus terrestres y C.
denticulata de la selva de Madre de Dios en Perú (Del Valle et al., 2018;
Esser et al.,2016; Rojas-Jaimes et al., 2021).

Figura 2. (a) observación directa
a 400X de vísceras y glándulas salivales de A. naponense colectado de P.
tajacu donde se observa levaduras. (b) cultivo vísceras y glándulas
salivales de A. naponense colectados de P. tajacu vistas al
microscopio a 400X donde se observa Fusarium sp.
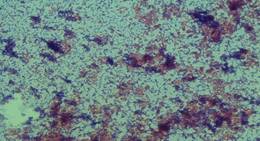
Figura 3. Observación
directa a 400X de cultivo de hígado de hámster retado con vísceras y
glándulas salivales de A. naponense colectado de P. tajacu donde
se observa levaduras.
En el presente
estudio no se logró aislar Leishmania en cultivo de intestinos y
glándulas salivales de las garrapatas ni se pudo identificar en la presencia
del parásito en los hámsteres retados con los inóculos de los tejidos de las
garrapatas, con excepción del control positivo con la
cepa JM de Leishmania. No obstante, Campos et
al. (2014) lograron causar infección por Leishmania en hámsteres
inoculados con extractos de garrapatas Rhipicephalus sanguineus
colectadas de perros. Si bien en el presente estudio no se identificó a Leishmania
en A. naponensis y A humerale, Araúz et al. (2016) identificaron
la forma de promastigote de Leishmania en glándulas salivales e
intestinos de R. sanguineus.
Araúz, M., Guerrero. F., Miranda de Oliveira, B., Costa de Aquino,
M., Hudson, S., Dias de Melo, G., Gomez, A., Takami, C., Garcia, M., Andreotti,
R., Félix de Lima, V. & Saraiva, K. (2016). Identification
of Leishmania spp. promastigotes in the intestines, ovaries, and salivary
glands of Rhipicephalus sanguineus actively infesting dogs. Parasitology
Research, 115, 3479–3484.
Barros-Battesti, D., Arzua, M., & Bechara, H. (2006).
Carrapatos de Importância Medico-Veterinaria da Região Neotropical: Um Guia
Ilustrado para Identificação de Espécies. 10ma edição. Sao Paulo: Butantan
Publicação. p. 223.
Campos, J., & Costa, F. (2014). Participation
of ticks in the infectious cycle of canine visceral leishmaniasis, in Teresina,
Piauí, Brazil. Rev. Inst. Med. Trop. Sao Paulo, 56(4),
297-300.
Colombo, F. A., Odorizzi, R. M., Laurenti, M. D., Galati, E. A.,
Canavez, F., & Pereira-Chioccola, V. L. (2011). Detection of
Leishmania (Leishmania) infantum RNA in fleas and ticks collected from
naturally infected dogs. Parasitology Research, 109, 267–274.
Couto, D. V., Hans Filho, G., Medeiros, M. Z., Vicari, C. F. S.,
Barbosa, A. B., & Takita, L. C. (2014). American
tegumentary leishmaniasis—a case of therapeutic challenge. An Bras Dermatol,
89, 974–976.
Dantas-Torres, F. (2009). Canine leishmaniosis in South America. Parasit
Vectors, 2(Suppl.1), 1-8.
Del Valle-Mendoza, J., Rojas-Jaimes, J., Vasquez-Achaya, F.,
Aguilar-Luis, M. A., Correa-Nunez, G., Silva-Caso, W., & et al. (2018).
Molecular identification of Bartonella bacilliformis in ticks collected from
two species of wild mammals in Madre de Dios: Peru. BMC Res Notes, 11,
405.
Esser, H., Foley, J., Bongers, F., Herre, E., Miller, M., Prins, H.,
& Jansen, P. (2016). Host body size and the diversity of tick assemblages
on Neotropical vertebrates. International Journal for
Parasitology: Parasites and Wildlife, 5(3), 295-304.
Marotta, C., Dos Santos, P., Cordeiro, M., Da S. Barros, J.,
Bell-Sakyi, L., & Fonseca, A. (2018). Trypanosoma amblyommi sp. nov.
(Protozoa: Kinetoplastida) isolated from Amblyomma brasiliense (Acari:
Ixodidae) ticks in Rio de Janeiro, Brazil. Parasitol Open, 4,
e2.
Rojas-Jaimes,
J., Correa-Núñez, G., Rojas-Palomino, N., & Cáceres-Rey, O. (2017).
Detección de Leishmania (V) guyanensis en ejemplares de Rhipicephalus
(Boophilus) microplus (Acari: Ixodidae) recolectados en
pecaríes de collar (Pecari tajacu). Biomédica, 37(Supl.2), 208-214.
Rojas-Jaimes,
J., Lindo-Seminario, D., Correa-Núñez, G., & Benoit, D. (2021). Characterization
of the bacterial microbiome of Rhipicephalus (Boophilus) microplus
collected from Pecari tajacu “Sajino” Madre de Dios, Peru. Scientific
Reports, 11, 6661.
Savoia, D. (2015). Recent updates and perspectives on
leishmaniasis. J Infect Dev Ctries, 9,
588–596.
Solano-Gallego, L., Rossi, L., Scroccaro, A., Montarsi, F., Caldin,
M., Furlanello, T. & et al. (2012). Detection of Leishmania
infantum DNA mainly in Rhipicephalus sanguineus male ticks removed from
dogs living in endemic areas of canine leishmaniosis. Parasit
Vectors, 5, 98.
Veland, N.,
Espinosa, D., Valencia, B., Ramos, A., Calderon, F., Arevalo, J., & et al. (2011).
Polymerase chain reaction detection of leishmania kDNA from the
urine of peruvian patients with cutaneous. Am J Trop Med Hyg, 84(4),
556-561.