Abejas de
las orquídeas como bioindicadores del estado de conservación de un bosque
Orchid bees
as bioindicators of the state of conservation of a forest
Bleysin Rojas1; Oliver Vásquez
1; Alonso Santos-Murgas2,
*; Rafael Cobos3; Iris Y. Gómez Robles4
1
Universidad de Panamá; Facultad de Ciencias
Naturales, Exactas y Tecnología; Centro Regional Universitario de Coclé, Penonomé,
Panamá.
2 Universidad de Panamá,
Facultad de Ciencias Naturales, Exactas y Tecnología, Departamento de Zoología,
Ciudad Universitaria Dr. Octavio Méndez Pereira, Panamá.
3 Universidad de
Pamplona, Norte de Santander, Colombia.
4 Universidad de
Panamá, Facultad de Ciencias Naturales, Exactas y Tecnología, Centro Regional
Universitario de Coclé, Penonomé, Panamá.
*Autor
corresponsal: santosmurgasa@gmail.com (A. Santos-Murgas).
ID ORCID de los autores
B.
Rojas:
 http://orcid.org/0000-0002-4548-1202 O.
Vásquez: http://orcid.org/0000-0002-3664-1520
http://orcid.org/0000-0002-4548-1202 O.
Vásquez: http://orcid.org/0000-0002-3664-1520
A.
Santos-Murgas: http://orcid.org/0000-0001-9339-486X R.
Cobos: http://orcid.org/0000-0002-7574-5168
I.
Y. Gómez Robles: http://orcid.org/0000-0002-5099-6366
RESUMEN
Los bioindicadores son organismos que determinan
el impacto ambiental de un ecosistema y se utilizan a los euglossinos por
responder a cambio ambientales, son de fácil observación, y su taxonomía es
bien conocida. El objetivo del estudio fue
evaluar calidad ambiental de un bosque a través de las abejas de las orquídeas
como bioindicadores. En esta investigación, se realizaron colectas a nivel de
sotobosque, por medio de “Bottle Traps”, y colectas manuales por método de
zigzag, en el cual se emplearon redes entomológicas, las abejas serán atraídas
con atrayentes químicos como: aceite de eucalipto, esencia de eugenol y esencia
de vainilla. Lo cual obtuvimos una abundancia bastante al alta con una captura
de 1024 individuos y una riqueza de 38 especies, el género que domino el
estudio fue ele genero Euglossa con 89,5%, seguido por el género Eulaema
con 9,6% y género Eufriesea con apenas un 0,8%, para el género Exaerete
que no se logra capturar individuos, La curva de completitud para el muestreo
general realizado permite observar una representatividad del 99%. En
conclusión, consideramos que la calidad ambiental en Cerro Llorón, a pesar de
las acciones antrópicas se encuentra bastante estable, porque encontramos un
abundancia y riqueza de especie bien elevada.
Palabras clave: Apidae; Euglossini; bioindicadores; Río Indio Nacimiento;
Cerro Llorón.
ABSTRACT
Bioindicators
are organisms that determine the environmental impact of an ecosystem and
euglossine are used for responding to environmental changes, they are easy to
observe, and their taxonomy is well known. The study's objective was to
evaluate a forest's environmental quality through orchid bees as bioindicators.
In this investigation, collections were made at the undergrowth level by means
of "Bottle Traps" and manual pools by zigzag method, in which entomological
networks were used; the bees will be attracted by chemical attractants such as
eucalyptus oil, the essence of eugenol and vanilla essence. We obtained a relatively
high abundance with capture of 1024 individuals, and a richness of 38 species,
the genus that dominated the study was the genus Euglossa with 89.5%, followed
by the genus Eulaema with 9.6% and the genus Eufriesea with just 0.8 %, for the
genus Exaerete that it is not possible to capture individuals, The completeness
curve for the general sampling carried out allows us to observe
representativeness of 99%. In conclusion, despite anthropic actions, the
environmental quality in Cerro Llorón is quite stable because we found a very
high abundance and species richness.
Keywords: Apidae; Euglossini; bioindicators; Rio Indio
Nacimiento; Cerro Lloron.
Recibido: 25-07-2022.
Aceptado: 22-09-2022.

INTRODUCCIÓN
Actualmente
una de las problemáticas mundiales es la pérdida de biodiversidad, debido a las
malas acciones de la población en general, como la tala indiscriminada, la
deforestación para construcción de viviendas y otras edificaciones, la emisión
de CO2 por los carros y fábricas, así como la mala disposición de
materiales contaminantes, y por supuesto la falta de políticas ambientales que
dan como resultado la poca conservación de los bosques, en donde se encuentran
especies esenciales para la polinización de orquídeas y de otras plantas de
suma importancia para el equilibrio de los ecosistemas (Meléndez et al., 2015;
Reyes-Novelo et al., 2009).
Según la
Autoridad del Canal de Panamá (ACP, 2017; 2018), en Río Indio Nacimiento, tramo
alto de la Cuenca de Río Indio, la ganadería extensiva, la agricultura de
subsistencia y en menor parte la agricultura comercial, son las actividades
mayormente realizadas por la población local, el mayor daño que sufre el
paisaje se da por las vías de acceso o los llamados “caminos de penetración”;
sin embargo, en el área de Cerro Llorón se puede observar la fragmentación del
bosque producida por la intervención antrópica.
Con respecto a
lo mencionado anteriormente surge el interés por los organismos bioindicadores,
de modo que se empleará Apidae: Euglossini como grupo bioindicador, ya que las
mismas son fácilmente observables, así como sensibles a perturbaciones
naturales, puesto que muy pocas especies pueden sobrevivir en áreas con cambios
en el uso del suelo (Ramírez et al., 2015). Además, es
importante mencionar que estas abejas son solitarias, y no producen miel, por
lo que su beneficio es la polinización, al contrario de las que producen miel
(Parra et al., 2016; Ramírez et al., 2015). Hay que resaltar que las abejas que
producen miel son las más utilizadas en el mundo para determinar el deterioro
del ambiente, en cambio las abejas de las orquídeas están siendo introducidas a
este campo de la investigación. Cabe señalar que para el área de estudio no
se han realizado con anterioridad monitoreos y/o muestreos con abejas de las
orquídeas, por lo que no hay documentación previa en cuanto a su diversidad y
abundancia.
El objetivo de
esta investigación es evaluar la calidad ambiental de un bosque en Cerro
Llorón, Rio Indio Nacimiento, Panamá Oeste, determinando la diversidad y
abundancia de las abejas de las orquídeas para comprender la panorámica
ambiental y aspectos ecosistémicos de estas abejas presentes en estos bosques.
Este estudio fue realizado
en Cerro Llorón, el cual está situado dentro de
una propiedad privada en Río Indio Nacimiento, Panamá Oeste. El sitio comprende
dos áreas de estudio, una zona intervenida (ZI), y una zona de bosque (ZB) (Tabla
1). Se llevaron a cabo dos muestreos por mes, por espacio de cuatro meses (mayo-septiembre
de 2021). La captura de las abejas se realizó mediante dos técnicas de
muestreo: método de trampas de botella (TB) y método de zigzag.
Se empleó un total de 8 TB para euglosinos,
elaboradas con botellas de 2,5L con un orificio de entrada (Vélez y Barrios,
2005; Razo, 2015; Reyes, 2020), en su interior constan de un atrayente químico
artificial, aceite de eucalipto para ambos sitios, instaladas a altura de
sotobosque (1.5 a 2 m), seleccionándose cuatro árboles al azar por sitio para
colgar las TB, siendo monitoreadas durante la mañana (11:00 a 11:30 am) en cada
zona.
Tabla 1
Información
de los sitios de muestreo
|
Sitios
de muestreo
|
Coordenadas
|
Altura
|
|
ZI
ZB
|
8º39’26,68”
N
-80º06’26,68”
W
8º39’48,94”
N
-80º06’42,61”
W
|
648
msnm
890
msnm
|
Como segundo método de
muestreo se estableció un sistema espacial, que consistió en un transepto de
tres diagonales (en zigzag) de hilo pabilo, de extremo a extremo de cada árbol,
a una altura aprox. de 1,70 a 2 m del suelo, en el mismo se colocaron tres atrayentes
(aceite de eucalipto, esencia de vainilla y eugenol) impregnados en algodón,
finalmente las abejas fueron colectadas con redes entomológicas (Osorio y
Santos, 2017; Santos y Añino, 2016; Santos et al., 2018; Machado et al., 2018).
Estas colectas se realizaron por un período de dos horas continuas.
Las
abejas colectadas se colocaron en viales con alcohol al 70%, para su posterior
transporte al laboratorio, en donde se procesaron e identificaron con la clave
taxonómica de Roubik & Hanson (2004). Adicional se tomaron datos de
variables abióticas para ambas áreas de muestreo: temperatura (°C) y humedad
relativa (%) (Machado et al., 2018; Vilhena et al., 2017).
Los datos recopilados,
fueron ordenados en una base de datos de Microsoft Excel 2016. Para cada área
del presente estudio, la completitud fue validada mediante el paquete R”iNext”
el cual se fundamenta en lo propuesto por Chao & Jost (2014), Colwell et
al. (2012) y Chao et al. (2015), el cual permite realizar las estimaciones para
muestras estandarizadas con un tamaño de muestra común o completa basado en los
números de Hill relacionando la riqueza (diversidad de orden cero) con la
abundancia (Hsieh et al., 2016).
Mediante el software “HillNumbersProfile”
de la autoría de Anne Chao se elaboraron los perfiles de diversidad, donde la
diversidad de 0D equivale al número de especies efectivas
halladas, ya que no contempla los valores de abundancia, la diversidad de 1D
representa el número de especies efectivas abundantes, por lo que se refiere a
la diversidad del estudio en un área específica, y la diversidad de 2D
representa al número efectivo de especies dominantes, ya que este es más
sensible a los valores representativos sin tomar los singleton y doubleton
(Jost, 2010).
Además, se elaboraron
curvas de rango abundancia en Excel 2016, para observar los patrones de
distribución de las especies de abejas colectadas en ZI y ZB (Carmona &
Carmona, 2013), y un análisis de similaridad de Jaccard en el software PAST (paleontological
statistics software) versión 4.08 para relacionar el número de especies compartidas
con el número total de especies exclusivas (Jaccard, 1908). Por último, se realizó
un análisis de correspondencia canónica donde se relacionaron las especies de euglosinas
con las variables abióticas tomadas y los sitios de muestreo en PAST.
En este estudio se
colectaron 1024 especímenes de abejas de la Tribu Euglossini distribuidos en
tres de los cuatro géneros reportados para Panamá (Roubik & Hanson, 2004),
siendo Euglossa Latreille (1802) el género con mayor riqueza de especies
(29), seguido de Eufriesea Cockerell (1908) (5 especies) y Eulaema Lepeletier
(1841)
(4
especies); asimismo, tenemos que el género Euglossa fue el más abundante
con 89,5% de la muestra total, estos resultados son similares a los obtenidos
en el estudio realizado en el Parque Nacional Darién por Santos et al. (2018).
De las especies del género Euglossa:
Euglossa hansoni (173 individuos), Euglossa dodsoni (150
individuos), Euglossa imperialis (74 individuos), Euglossa championi
(71 individuos) y Euglossa tridentata (70 individuos) fueron las más
abundantes (Figura 1), en general este género es el más rico en especies
comprendiendo más de 140, las cuales se encuentra subdivididas en siete
subgéneros (Ferrari et al., 2017). Además, Chilito-Rodríguez et al. (2021)
explican que “Las especies pertenecientes a este género suelen ser genera-listas
y presentan la capacidad de adaptarse a los cambios en las coberturas
vegetales”.

Figura 1. Géneros más abundantes durante el muestreo realizado en Río Indio. A)
Género Euglossa. B) Género Eufriesea. C) Género Eulaema.
El género Exaerete se
caracteriza por ser abejas cleptoparásitas de otros dos géneros de Euglossini
(Roubik & Hanson, 2004; Oliveira, 2011) y además presentan cierta
preferencia por zonas con vegetación densa, por lo que comúnmente son
colectadas en bosques de galería (Chilito-Rodríguez et al., 2021), sin embargo,
tomando esto en cuenta, para este estudio no se logró capturar ningún miembro
para este grupo. Por otra parte, se sabe que estas abejas no son comunes en los
muestreos (Chilito-Rodríguez et al., 2021).
El estimador de completitud
del muestreo para cada método de colecta usado, registra valores entre el 98,53
% y 99,27%, lo cual da soporte estadístico a los resultados; así mismo, la
curva de rarefacción muestra una estabilización del eje de la riqueza y el intervalo
de confianza (sombra) en ambos métodos permite inferir que con una mayor
densidad del muestreo la riqueza podría aumentar, sin embargo, teniendo en
cuenta el grado de completitud, dichas especies serían consideradas raras y no
definirían la comunidad (Tabla 2 y Figura 2).
Tabla 2
Integridad
del muestreo
|
|
Trampa
de botella
|
Zigzag
|
|
Abundancia
Riqueza
observada
Representatividad
|
337
26
0,9853
|
687
37
0,9927
|
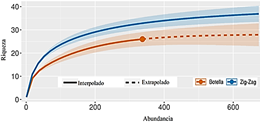
Figura 2. Curva de completitud basada en rarefacción
de las especies de abejas euglosinas colectadas por método de zigzag y Trampa
de botella mediante el método Chao & Jost (2012).
Según Grandolfo et al.
(2013) la diferencia presente en la riqueza y abundancia en diferentes trabajos
pueden ser variable debido a diversos factores como el esfuerzo de muestreo, la
estacionalidad, el clima, y el área de muestreo entre otros factores que
influyen en la cantidad de especies colectadas, sin embargo, el uso de varios
métodos de muestreo combinados permite la obtención de un mejor registro de
especies, por lo que esto complementa, y amplia la muestra (Oliveira-Junior et
al., 2015; Pardo & Gonzalez, 2007). Según Parra et al., 2016
dijo que los métodos de trampas de botellas y colecta activa por medio de redes
entomológicas son complementarios para varios sitios simultáneos.
El uso de más de un método
de colecta permite tener una mejor consideración de la riqueza y representatividad
de las poblaciones, ya que, de emplear un solo método, la riqueza de abejas
sería más baja de lo esperado, tal y como se muestra en el estudio realizado
por Cordeiro et al. (2019), en donde al solo muestrear con trampas el estimador
mostró un 60-89% de abejas colectadas. Además, Nemésio & Vasconcelos
(2014), sugieren que a la hora de realizar estudios siempre se deben considerar
una buena metodología que brinde una validez científica.
Los índices clásicos
empleados para determinar la diversidad indican lo siguiente: en el índice de
dominancia del inverso de Simpson_1-D para las trampas de botella (TB) fue de
0.8894 y zigzag fue de 0.9224, siendo el método de zigzag el que cuenta con una
mayor probabilidad (3.3%) de que los dos individuos tomados al azar en la
muestra no sean iguales. El índice de Shannon (H) se obtuvo 2.504 para TB, y
2.912 para zigzag, mostrando que este último es levemente más diverso, entrando
dentro del rango normal; en cuanto al índice de Margalef el método de zigzag
arroja un valor de 5.511 y de 4.295 para el método TB, por lo que, el área
muestreada presenta alta diversidad, siendo mejor representada por el método de
Zigzag. Por último, el índice de Pielou (Equitatibility_J) muestra que la
abundancia de las especies colectadas por método de zigzag es más homogénea con
respecto a la obtenida por TB. Por ende, estos resultados determinan que el
método de zigzag presenta una mayor colecta de especies, y guardan cierto gado
de similitud con los obtenidos por Nemésio et al., (2014), aun así,
exponen el hecho de que estos datos no deberían ser comparados con los métodos
para los índices clásicos de diversidad, sino que más bien deben evaluarse con
un método analítico que contemple la identidad y abundancia de las especies. Para los
biólogos el estudio de la biodiversidad es sumamente importante, por lo que el
concepto de diversidad es bastante amplio, y a lo largo de los años va tomando
relevancia en la ecología de comunidades como en la biología de conservación,
no obstante, su definición es tan variada que dificulta su entendimiento, a la
vez que las herramientas matemáticas empleadas para su medición no siempre
resultan ser las adecuadas, lo que conlleva a una gran polémica, sin embargo
este concepto ha ido evolucionando de ser un mero recuento de especies a
cálculos más complejos que contemplan las abundancias relativas y el tiempo de
divergencia evolutiva entre especies (Moreno et al., 2011; Jost &
González-Oreja, 2012; Jost, 2018). Es por lo que
considerando lo anteriormente descrito, se estimó la diversidad en números efectivos
de especies, también conocido como números de Hill o índices de diversidad
verdadera, los cuales, según Jost (2018) son equivalentes a transformaciones de
los índices de diversidad tradicionales en unidades más coherentes con la
misam, facilitando la interpretación, y, además, obedecen al principio de
replicación.
Con base en lo expresado
anteriormente, la diversidad verdadera estimada para TB de orden cero (0D)
equivalente a la riqueza es 26 especies efectivas, en el orden uno 1D
= 12 (especies efectivas abundantes) y orden dos, 2D = 9
(especies efectivas dominantes); en cuanto al método de zigzag, 0D
es de 37 especies efectivas, de las cuales 18 especies son abundantes (1D)
y 13 especies dominan (2D), dado que la riqueza (0D)
no contempla la abundancia y la dominancia (2D) no tiene en cuenta
la abundancia de las especies raras, se realiza la comparación de diversidad
usando el orden uno, donde el método de Zigzag es 33.53% más diverso que el
método de trampas de botella (Figura 3). Estos resultados concuerdan con los
estudios realizados en el Parque Nacional Cusuco, Honduras (McCravy et al.,
2016), y con los obtenidos por Nemésio et al., (2014), así como los de
Oliveira-Junior et al., (2015), en donde la metodología con redes obtuvo
una significativa colecta de especies, además indican que esté método puede
revelar la composición faunística real de los sitios de muestreo, sin embargo
hay que considerar el hecho de que está metodología está restringida para
estratos de sotobosque (Pardo et al., 2007), por lo que la metodología
de trampa no debe ser descartable, además está suele ser recomendada para
muestreos complejos que requieran replicaciones, y para muestrear en diferentes
estratos del bosque (Sydney & Gonçalves, 2015; Reyes Ledezma et al. 2020),
en contraste Oliveira-Junior et al. (2015) recomienda el uso de múltiples
métodos que amplíen la muestra de la fauna.
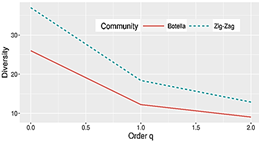
Figura 3. Perfiles de diversidad de las abejas de las orquídeas de Cerro Llorón,
Río Indio Nacimiento.
Las curvas de
rango-abundancia muestran que la composición y abundancia entre especies del
sitio de muestreo difieren un poco de acuerdo con el método de colecta y la
zona muestreada, por ende, se tiene que las especies dominantes para el área
intervenida por el método de zigzag fueron Eg. hansoni y Eg. dodsoni,
en cambio para TB fueron Euglossa gorgonensis y Eg. dodsoni. Por
otro lado, para el área boscosa con el método de zigzag las especies dominantes
fueron Eg. dodsoni y Eg. championi, sin embargo, por el método de
TB las más dominantes serian Eg. hansoni, seguida de Eg. dodsoni.
En general, se observa que las demás especies mantienen cierta estabilidad. Por
otra parte, Carneiro et al. (2018); Andrade-Silva et al. (2012), también
mencionan que el cambio de una vegetación cerrada a una más abierta puede
influir en el ensamblaje de las abejas de las orquídeas, esto puede
considerarse para las especies que no presentaron colecta en una u otra área.
Otro factor importante es la movilidad que presentan estas abejas, recordando
aquí que las mismas se mueven en busca de recursos alimenticios o de material
para sus nidos (Santos et ál. 2018; Ferreira et al., 2013), en nuestro estudio
las dos áreas de muestreo estuvieron separadas por una distancia aproximada de
2km, creemos que esto pudo haber influido entre el flujo de individuos.
Con relación a Eg.
hansoni y Eg. dodsoni, se sabe que ambas son abejas pequeñas (10mm
de largo), y de lengua corta (Roubik & Hanson, 2004; McDonald et al.,
2022), la primera presentó una abundancia de 106 individuos (repartidos de la
siguiente manera: ZB=41, ZI = 65) para el método de zigzag, y 67 individuos
(repartidos en ZB= 56, ZI= 11) para TB, para Armas (2009) esta abeja es
sensible a las perturbaciones, puesto que se ha encontrado en reservas y áreas
protegidas que presentan ciertas características climáticas, sin embargo, en
nuestro estudio podemos ver claramente que en área boscosa mantuvo estabilidad,
pero la mayor dominancia de acuerdo al método de zigzag fue para el área
intervenida. En cuanto a la segunda, se puede decir que su abundancia por los
dos métodos de colecta se mantuvo estable, sin embargo, se observa que tuvo
mayor predominancia en el área boscosa, por lo que coincide con lo dicho por
Riveros et al. (2009), quienes mencionan que la misma es propia de bosques
nubosos de Costa Rica y Panamá, así mismo, también describen que prefieren
zonas con árboles altos que les brinden protección a sus nidos contra vientos
fuertes, no obstante, varios aspectos de su biología son desconocidos.
Eg. gorgonensis, colectada en
mayor cantidad por trampa de botella, con una totalidad de 27 individuos, fue
la más abundante para el área intervenida con 23 especímenes colectados, esta
especie exhibe una gran variación de color dependiendo de su rango de
distribución, lo cual influye en su identificación (Hinojosa-Díaz & Brosi,
2013).
 Eg. championi presentó con el método de zigzag una
abundancia relativamente alta (46 individuos), con una preferencia por ZB, en
contraste para TB su abundancia es baja con solo un individuo muestreado para
cada área; está abeja tiene un tamaño aproximado de 13 mm de largo, de lengua
corta (Roubik & Hanson, 2004).
Eg. championi presentó con el método de zigzag una
abundancia relativamente alta (46 individuos), con una preferencia por ZB, en
contraste para TB su abundancia es baja con solo un individuo muestreado para
cada área; está abeja tiene un tamaño aproximado de 13 mm de largo, de lengua
corta (Roubik & Hanson, 2004).
La similitud entre la fauna
de euglosinas evidencia que, al comparar las áreas de muestreo, por método de
zigzag estas comparten aproximadamente un 70% de las especies colectadas en
ambas comunidades, mientras que un 30% son abejas que no se comparten entre
ambos sitios de muestreo, por tanto, en trampas de botellas es más bajo la
similaridad que comparten solo un 45% aproximadamente, lo restante son especies
diferentes.
El análisis de
correspondencia canónica para las especies de abejas euglossinas en relación
con las variables climáticas de humedad y temperatura, expone que estás fueron
determinantes para la distribución de las abejas, coincidiendo con Andrade-
Silva et al. (2012). Según Vilhena et al. (2017), los machos de la tribu
euglossini se muestran más activos cuando la temperatura oscila entre 24.5 °C a
27 °C, y cuando la humedad relativa es superior al 100% podría ser adversa para
abejas pequeñas como lo son las del género Euglossa que presentan una
menor capacidad de termorregulación, ejemplificando este hecho, podemos decir
que durante los muestreos al haber un aumento en la humedad relativa, y una
baja temperatura, inmediatamente cesaba la actividad o el flujo de euglosinos,
y en el instante en que la temperatura se regulaba volvían a acercarse a los
cebos. De acuerdo con Machado et al. (2018) la variable de humedad relativa
puede explicar las diferencias en las comunidades.
Los
resultados del presente estudio permiten expresar que pese a la intervención
antrópica en Cerro Llorón, la diversidad de abejas de las orquídeas que alberga
es alta, lo cual indica el grado de conservación del área muestreada; por lo
tanto, es un área a tener presente en planes de conservación y ecoturismo
beneficiando a los pobladores de Rio Indio, cuya actividad al momento del
presente estudio, no ha incidido negativamente sobre la biodiversidad de la
cobertura vegetal conservada, para lograr esto, debemos incentivar y educar a
la población sobre la conservación, así como contar con el apoyo de las
respectivas autoridades. Además, brindar a los lugareños las herramientas
necesarias para evitar que el tipo de agricultura practicada no afecte a estos
organismos, pues son necesarios para mantener el equilibrio en estos
ecosistemas de suma importancia para la humanidad.
El
género con mayor captura de individuos fue Euglossa con 918 individuos y
con 5 especies más abundantes, entre ellas tenemos Eg. hansoni, Eg. dodsoni,
Eg. imperialis, Eg. championi y Eg. gorgonensis.
Para
este estudio el método de zigzag fue el más efectivo durante la captura de
estas abejas, aunque sugerimos el empleo de otros métodos de muestreo
complementarios, para obtener unos mejores resultados. En cuanto a la similitud
de las comunidades de abejas, para ambas áreas por el método de zigzag, esta
fue bastante alta indicando que son muy similares, sin embargo, en cuanto a la
similitud por el método de TB, la similaridad es más baja indicando que la
mayoría de las especies son diferentes.
También
es importante tomar en cuenta las varia-bles ambientales, porque estas son
imprescindibles para entender los patrones de distribución de las abejas de las
orquídeas, así como el de comprender sus preferencias de hábitat.
Por
último el estudio sobre la diversidad y abundancia de abejas de las orquídeas
en bosques de tierras altas contribuye en gran medida al conocimiento de la
distribución biogeográfica de las mismas, así como el estado en el que se
encuentran estos sitios poco estudiados, no obstante es necesario ampliar este
estudio pues en mayor medida fue realizado durante la estación lluviosa, por lo
que se necesita recopilar información de la estación seca que permita complementar
la información obtenida.
A
la familia Gil Espinosa, en especial al Sr. Samuel Gil, quién nos concedió el
permiso para realizar este trabajo, así como nos brindó de su guía a lo largo
de todo lo que fue el desarrollo de este estudio. También a todos nuestros
familiares, y amigos que estuvieron con nosotros en cada paso apoyándonos, y en
especial a los profesores quienes con mucha paciencia nos aconsejaron y
brindaron de todos sus conocimientos.
REFERENCIAS
BIBLIOGRÁFICAS
Autoridad del Canal de Panamá. (2017).
Línea base socioeconómica general. Cuenca de Río Indio. Objetivo 4. Tarea 17a.
Informe de línea base socioeconómica general de la cuenca de río indio.
Autoridad del Canal de Panamá. (2018).
Programa de monitoreo y conservación cuenca de Río Indio. Objetivo 2. Tarea 9d.
Evaluación de la biodiversidad en la cuenca de Río Indio y diseño de programa
de monitoreo y conservación (Año 2).
Andrade-Silva, A. C. R., Nemesio, A.,
Oliveira, F. F. y Nascimento, F. S. (2012). Spatial-Temporal
Variation in Orchid bee communities (Hymenoptera: Apidae) in remnants of
Arboreal Caatinga in the Chapada Diamantina Region, State of Bahia, Brazil. Neotropical
Entomology, 41, 296-305.
Armas, Q. A.G. (2009). Riqueza y
Distribución Potencial de las Abejas Euglosinas (Apinae: Euglossini) en
Guatemala (Tesis de Licenciatura). Universidad de San Carlos de Guatemala,
Guatemala, Guatemala.
Carmona, V., & Carmona, T. (2013).
La diversidad de los análisis de diversidad. Bioma, 14,
20-28.
Carneiro, L. S., Aguiar, W. M., &
Santos, G. M. M. (2018). Orchid bees (Hymenoptera: Apidae:
Euglossini) in Seasonally Dry Tropical Forest (Caatinga) in Brazil. Sociobiology,
65(2), 253-258.
Chao, A., Chiu, C. H., & Jost, L.
(2014). Unifying species diversity, phylogenetic diversity, functional
diversity, and related similarity and differentiation measures through Hill
numbers. Annual Review of Ecology, Evolution, and Systematics, 45(1),
297–324.
Chao, A., Jost, L., Hsieh, T. C., Ma,
K. H., Sherwin, W., & Rollins, L. A. (2015). Expected Shannon entropy and
Shannon differentiation between subpopulations for neutral genes under the
finite island model. PLOS ONE, 10(6), e0125471.
Chilito-Rodríguez, E. P., Bañol, E. R. H.,
Valencia, A. V., & Parra Olarte, M. J. (2021). Abejas Euglossinas (Apidae:
Euglossini): un enfoque de conservación e importancia ecológica. Brazilian
Journal and Enviromental Research, 4(4), 6224-6245.
Colwell, R.K., Chao, A., Gotelli,
N.J., Lin, S.‐Y., Mao, C.X., Chazdon, R.L., & Longino, J.T. (2012).
Models and estimators linking individual‐based and sample‐based
rarefaction, extrapola-tion and comparison of assemblages. Journal of Plant
Ecology, 5, 3–21.
Cordeiro, M., Garrafoni, A. R. S.,
& Lourenço, A. P. (2019). Rapid assesment of the orchid bee fauna
(Hymenoptera: Apidae: Euglossini) in the vicinity of an urban Atlantic Forest
remnant in Sao Paulo, Brazil. Brazilian Journal of Biology, 79(1),
149-151.
Ferrari, R. R., Nemésio, A. &
Silveira, F.A. (2017). Phylogeny of the orchid-bee by genus Euglossa Latreille
(Hymenoptera: Apidae), with emphasis on the subgenera E. (Glossura) Cockerell
and E. (Glossorupoda) Moure. Zoologischer Anzeiger- A Journal of Comparative
Zoology, 267, 82-100.
Freiria, G. A., Garófalo, C. A., &
Del Lama, M. A. (2017). The primitively social behavior of Euglossa cordata
(Hymenoptera, Apidae, Euglossini): a view from the perspective of kin selection
theory and models of reproductive skew. Apidologie, 48,
523–532.
Grandolfo, V. A., Bozza Junior, R. C.,
Silva Neto, C. M., Neto, J. N.M., & Gonçalves, R. B. (2013). Riqueza e
Abundância de abelhas Euglossini (Hymenoptera, Apidae) en Parques Urbanos de
boiânia, Goiás. Entomo Brasilis, 6(2), 126-131.
Hinojosa-Díaz, I., & Brosi, B. J.
(2013). First records and description of metallic red females of Euglossa
(Alloglossura) gorgonensis Cheesman, with notes on color variation within the
especies (Hymenoptera, Apidae). Zookeys, 335, 113-119.
Hsieh, T. C., Ma, K. H., & Chao,
A. (2016) iNEXT: An R package for interpolation and extrapolation of species
diversity (Hill numbers). Methods in Ecology and Evolution, 7,
1451-1456.
Jaccard, Paul. (1908). Nouvelles
Recherches Sur la Distribution Florale. Bulletin de la
Societe Vaudoise des Sciences Naturelles, 44, 223-70.
Jost, L. (2010). The
relation between evenness and diversity. Diversity, 2(2),
207–232.
Jost, L. (2018). ¿Que
entendemos por diversidad? El camino hacia la cuantificación. Metodo
Science Studies Journal, 9, 55-61.
Jost, L., DeVries, P. J., Walla, T.,
Greeney, H., Chao, A., & Ricotta, C. (2010). Partitioning diversity for
conservation analyses. Diversity and Distributions, 16(1), 65–76.
Jost, L., & González-Oreja, J. A.
(2012). Midiendo la diversidad biológica: más allá del índice de Shannon. Acta
zoológica lilloana, 56 (1-2), 3-14.
McCravy, K. W., Dyke, J. V., Creedy,
T. J. & Roubik, D. W. (2016). Orchid bees (Hymenoptera: Apidae: Euglossini)
of Cusuco National Park, State of Cortés, Honduras. Florida
Entomologist, 99(4), 765-768.
McDonald, E., Podesta, J., Fortuin, C.
C., & Gandhi, K. J. K. (2022). Expanded distribution range of
nine orchid be especies (Hymenoptera, Apidae, Euglossini) in Costa Rica. Preprints
ARPHA.
Meléndez, R. V., Ayala, R., &
Delfín, G. H. (2015). Abejas como bioindicadores de perturbaciones en los
ecosistemas y el ambiente. En C. A. González. Zuarth, A. Vallarino, J. C.
Pérez. Jiménez, y A. M. Low. Pfeng (Eds.), Bioindicadores: Guardianes de nuestro
futuro ambiental (pp. 349-372). México D. F., México: El Colegio de la Frontera
Sur (ECOSUR) e Instituto Nacional de Ecología y Cambio Climático (INECC).
Nemésio, A., & Vasconcelos, H. L.
(2014). Effectiveness of two sampling protocols to survey orchid bees
(Hymenoptera: Apidae) in the Neotropics. J Insect
Conserv, 18, 197-202.
Oliveira-Junior, J. M. B., Almeida,
S. M., Rodrigues, L., Silvério Junior, A. J., & Anjos-Silva, E. J. (2015). Orchid
bees (Apidae: Euglossini) in a forest fragment in the ecotone Cerrado-Amazonian
Forest, Brazil. Acta Biológica Colombiana, 20(3),
67-78.
Osorio, A. M., & Santos, M. A.
(2017). Estudio sobre el conocimiento de la diversidad de abejas de la Orquídea
(Apidae: Euglossini) en la comunidad de Ustupu, Comarca Kuna Yala, Panamá. Scientia
(Panamá), 28(1), 61-71.
Padrón, P. S., Roubik, D. W., &
Picón, R. P. (2018). A Preliminary Checklist of the Orchid Bees
(Hymenoptera: Apidae: Euglossini) of Ecuador (Review). Hindawi
Psyche, (2018), Article ID 2678632.
Pardo, A. S. & Gonzalez, V. H.
(2007). Diversidad de Abejas (Hymenoptera: Apoidea) en estados sucesionales del
bosque húmedo tropical. Acta Biológica Colombiana, 12(1), 43-56.
Parra, A. H., Tupac, J. O., Sandino,
J., & Ospina, R. T. (2016). Abejas de la orquídea (Hymenoptera: Apidae:
Euglossini) y su importancia como polinizadoras de amplio rango en ecosistemas
naturales. En G. Nates (Ed.), Iniciativas colombianas de polinizadores (pp.
141-155). Colombiano, Bogotá: publicado por la Universidad Nacional Colombia
F.C.N. Departamento de Biología.
Razo, L. A. E. (2015). Abejas
silvestres (Hymenoptera: Apoidea: Anthophila) y sus interacciones con la flora
en la Sierra de Quila, Telocotlán, Jalisco (Tesis de maestría). Universidad
de Guadalajara, Jalisco, México.
R Core Team (2019). R: A language and
environment for statistical computing. R Foundation for Statistical Computing,
Vienna, Austria. URL https://www.R-project.org/
Reyes-Novelo, E., Meléndez R., V.,
Delfín, G. H., & Ayala, R. (2009). Abejas silvestres como bioindicadores en
el neotrópico. Agroecosistemas Tropicales y Subtropicales, 10(1), 1-13.
Reyes Ledezma, K. Y., Santos Murgas,
A., González, P., Gómez, I. Y. y Vargas, A. B. (2020). Diversidad alpha y beta
de abejas euglossini (Hymenoptera: Apidae) en el dosel y sotobosque del Cerro
Turega, Provincia de Coclé, Panamá. Tecnociencia, 22(2), 205-225.
Riveros, A. J., Hernández, E. J., & Wcislo, W.
T. (2009). Nesting Biology of Euglossa dodsoni Moure (Hymenoptera:
Euglossinae) in Panama. Journal of the Kansas Entomological Society, 82(2),
210-214.
Roubik, D. W., & Hanson, P. E.
(2004). Orchid bees of tropical America: Biology and field guide. Santo
Domingo de Heredia, Costa Rica: Instituto Nacional de Biodiversidad (InBio).
370 pp.
Santos, M. A., & Añino, R. Y. J.
(2016). Contribución al conocimiento de la diversidad de abejas de las
orquídeas (Apidae: Euglossini) de la Península de Azuero, Panamá. Tecnociencia,
18(2), 45-58.
Santos, M. A. (7/2018). Abejas de las
Orquídeas (euglossinae) como indicadora de biodiversidad en los bosques
tropicales y salud de los ecosistemas. {presentación de diapositiva}.
Santos, M. A., Abrego, J. C., López
Ch., O. G, Monteza, C., Osorio, M., Guardia, R., …Villarreal, C. (2018). Abejas
de las Orquídeas (Hymenoptera: Apidae: Euglossini) del Parque Nacional Darién,
Panamá. Tecnociencia, 20(2), 59-69.
Sydney, N. V., & Gonçalves, R. B.
(2015). Is the capture success of orchid bees (Hymenoptera, Apoidea) influenced
by different baited trap designs? A case study from southern
Brazil. Revista Brasileira de Entomologia, 59, 32-36.
Vilhena, P. S., Rocha, L. I., &
Garófalo, C. A. (2017). Male orchid bees (Hymenoptera: Apidae:
Euglossini) in canopy and under-story of Amazon Várzea Floodplain Forest. I.
Microclimatic, Seasonal and Faunal Aspects. Sociobiology, 64(2),
191-201.