 |
INTRODUCCIÓN
Riqueza de especies de mariposas (Lepidoptera: Papilionoidea) en un área de bosque tropical en Zarumilla, Tumbes, Perú
Species Richness of butterflies (Lepidoptera: Papilionoidea) in a tropical forest area in Zarumilla, Tumbes, Peru
Euder J. Juárez-Nima1, *; Pedro S. Castillo-Carrillo2; Gerardo Lamas3
1 Asociación Regional de Productores de Cacao Tumbes, km 7 Carretera a Papayal, Villa Uña de Gato, Tumbes, Perú.
2 Universidad Nacional de Tumbes. Ciudad Universitaria, Av. Universitaria S/N, Tumbes, Perú.
3 Departamento de Entomología, Museo de Historia Natural, Universidad Nacional Mayor de San Marcos, Apartdo14-0434, Lima-14, Perú.
* Autor corresponsal: javierjuarez2812@gmail.com (E. J. Juárez -Nima).
ORCID de los autores:
E. J. Juárez-Nima: https://orcid.org/0000-0002-7330-2910 P. S. Castillo-Carrillo: https://orcid.org/0000-0002-0255-1047
G. Lamas: https://orcid.org/0000-0002-3664-6730
RESUMEN
Las mariposas son indicadores del bienestar ambiental de los bosques y también, símbolo de belleza, colorido y emociones en los seres humanos. El conocimiento de este grupo en el área es parcial, razón por lo cual se desarrolló el estudio, desde mayo del 2020 hasta abril del 2021 en siete sectores del valle de Zarumilla, Tumbes, Perú. El objetivo fue evaluar la diversidad de mariposas a nivel de familias, utilizando los índices de Shannon (H) y Simpson (ʎ), su abundancia relativa y riqueza de especies en cada sector. Las evaluaciones fueron quincenales usando red entomológica y trampas Van Someren - Rydon y como cebo atrayente una mezcla de frutas, chicha de jora y melaza o azúcar en proporciones de 70, 20 y 10%; además de cebos a base de pescado en descomposición. Los ejemplares recolectados se depositaron en sobres de papel para su montaje e identificación. Se recolectó un total de 323 ejemplares distribuidos en seis familias: Nymphalidae (39,13%), Hesperiidae (27,54%), Pieridae (15,94%), Lycaenidae (10,14%), Papilionidae (2,90%) y Riodinidae (4,35%). Se obtuvo una riqueza de 69 especies distribuidas en: Hesperiidae (19), Lycaenidae (7), Nymphalidae (27), Papilionidae (2), Pieridae (11) y Riodinidae (3). Según el índice de Shannon (H), la mayor diversidad se encontró en el sector Uña de Gato. El índice de diversidad de Simpson (ʎ) fue alto en los sectores de Matapalo, Uña de Gato, Papayal, Lechugal, Manglares y La Coja.
Palabras claves. Índice de Shannon; Índice de Simpson; Manglares; Nymphalidae; Riodinidae.
ABSTRACT
Butterflies are indicators of the environmental well-being of forests and a symbol of beauty, color and emotions in human beings. The knowledge of this group in the area is partial, which is why the study was carried out from May 2020 to April 2021 in seven sectors of the Zarumilla Valley, Tumbes, Peru. The objective was to evaluate the diversity of butterflies at the family level, using the Shannon (H) and Simpson (ʎ) indices, their relative abundance and species richness in each sector. The evaluations were biweekly using entomological nets and Van Someren - Rydon traps and as an attractive bait a mixture of fruits, chicha de jora and molasses or sugar in proportions of 70, 20 and 10%; in addition to baits based on decomposing fish. The collected specimens were placed in paper envelopes for assembly and identification. A total of 323 specimens were collected distributed in six families: Nymphalidae (39.13%), Hesperiidae (27.54%), Pieridae (15.94%), Lycaenidae (10.14%), Papilionidae (2.90%) and Riodinidae (4.35%). A richness of 69 species was obtained distributed in: Hesperiidae (19), Lycaenidae (7), Nymphalidae (27), Papilionidae (2), Pieridae (11) and Riodinidae (3). According to the Shannon index (H), the greatest diversity was found in the Uña de Gato sector. The Simpson diversity index (ʎ) was high in the sectors of Matapalo, Uña de Gato, Papayal, Lechugal, Manglares and La Coja.
Key words. Index of Shannon; Indix Simpson; Mangroves; Nymphalidae; Riodinidae.
Aceptado: 09-08-2024
|
El departamento de Tumbes por su ubicación geográfica es diverso en fauna y flora. Cuenta con áreas naturales protegidas como El Parque Nacional Cerros de Amotape, El Santuario Nacional Los Manglares de Tumbes, El Área de conservación Ambiental Manglares delta de río Tumbes- Bahía de Puerto Pizarro, La Reserva Nacional de Tumbes, el Área de Conservación Angostura-Faical y el Área de Conservación Privada Bosque Seco Amotape (SERNANP 2019). Su importancia radica en albergar una entomofauna diversa y valiosa que puede ser aprovechada en la implementación de ideas de bionegocios con manejo sustentable y conservación.
Entre esta entomofauna, se encuentran las mariposas que son muy importantes, debido a que son consideradas como organismos indicadores de diversidad del estado de conservación de los ecosistemas y de los cambios de especies que ocurren como consecuencia del uso de suelo (Blair, 1999; Brown & Freitas, 2000; DeVries et al., 1999; Van Swaay et al., 2015; Comay et al., 2021); por la relación directa que presentan con sus plantas hospedantes (Wahlberg, 2001; Bonebrake et al., 2010); a su respuesta ante los cambios y perturbaciones ambientales; por ser sensibles a factores abióticos como la humedad, radiación solar y temperatura (Brown, 1997; Brown & Freitas, 2000; Kremen et al., 1993; Ospina & Reinoso, 2009); a su papel como proveedores de servicios ecosistémicos como la polinización (Gilbert, 1972; Tobar et al., 2001) y a la facilidad de hacerles seguimiento, colectar, manipular e identificar en condiciones de campo (Brown & Freitas, 2000; DeVries et al., 1997; DeVries, 1999), por ejemplo el Plan de Seguimiento de Mariposas en Europa es la red de monitoreo de invertebrados más grande del mundo, fundamentada principal-mente en la recolección de datos por voluntarios a lo largo de miles de transectos lineales (Sevilleja et al., 2020).
Por otro lado, son un grupo taxonómico muy estudiado a nivel mundial y son uno de los grupos de insectos más conocidos con diversos valores ecológicos, incluidos los polinizadores, ornamen-tales, culturales y científicos (Andrade-Correa, 1998; Kremen et al., 1993, Ospina & Reinoso, 2009; Wang et al., 2020). Adicionalmente, los estados adultos de las mariposas tienden a concentrarse en áreas donde las larvas pueden alimentarse de sus plantas hospederas (DeVries & Walla, 2001; Willmott & Freitas, 2006; Vásquez et al., 2017), por lo cual han sido propuestas como buenas indi-cadoras de la calidad del hábitat y, frecuentemente se utilizan en investigaciones ecológicas (Andrade-Correa, 1998; Warren et al., 2021). Así mismo, existe una estrecha relación desde la evolución de los lepidópteros y la de los vegetales debido a que han seguido caminos paralelos producto de los fenómenos de simbiosis que existen entre ambos y que las han llevado a una coevolución, estos insectos que representan apenas el 11% de todas las especies animales conocidas en el planeta (Andrade-Correa & Amat-García, 2007), se alimen-tan principalmente del néctar de las flores, mien-tras que los vegetales utilizan a las mariposas para favorecer su polinización (Sánchez-Tlacuahuac, 2023). Esta estrecha relación llega a tal extremo que con la alteración de un ecosistema, plantas y mariposas pueden seguir la misma suerte desapa-reciendo una si lo hace la otra (Medina, 2009). Las mariposas al igual que los abejorros son grupos funcionales clave con diferentes relaciones coevo-lutivas con las plantas (Alanen et al., 2011; Garrido et al., 2022).
Tienen propiedades específicas adicionales que las califican como un taxón complementario adecuado para las evaluaciones de conservación de la biodiversidad. Están bien estudiadas en compa-ración con otros grupos taxonómicos (Boggs et al., 2003), han demostrado tener una respuesta rápida a los cambios ambientales (Goulson et al., 2005; Thomas et al., 2004; Comay et al., 2021) y pueden usarse como especies paraguas para otros taxones de insectos (Thomas, 2005). Igualmente, las mariposas desempeñan un papel importante en los paisajes agroforestales, principalmente para realizar servicios ecosistémicos esenciales, a saber, la polinización y el mantenimiento de las relaciones presa-depredador (Keerthika & Parthiban, 2022).
En el ámbito de la ingeniería, se ha destacado la relevancia de estudiar las características de sus alas y la complejidad de la disposición de sus escamas. Estas alas presentan propiedades como flexibilidad, impermeabilidad, resistencia, ligereza, autolimpieza, suavidad y capacidad de termorregulación para la disipación y/o absorción del calor, entre otras. Específicamente, las especies Anarthia amathea (Linneaus, 1758) y Leptophobia aripa (Boisduval, 1836) han sido objeto de análisis mediante técnicas de caracterización como la microscopía de barrido electrónico y la fuerza atómica. Estos estudios han permitido diseñar prototipos para ecodiseño y bioconstrucción (Gasca et al., 2015).
Shuey et al. (2024), para generar información sobre la naturaleza de las comunidades de mariposas en Belice y las posibles estrategias para su conser-vación, analizaron un conjunto de datos de 31 539 ocurrencias que representan 1 047 especies de mariposas, encontrando que la mayoría de las especies en Belice rara vez se encuentran y el 23% se conocen de una o dos apariciones. Por el contrario, unas pocas especies (2%) son despro-porcionadamente abundantes (19% de las apariciones) y que la mayoría de las 'especies desa-parecidas' pertenecen a las familias Lycaenidae, Riodinidae y Hesperiidae, grupos difíciles de identificar y que tienen rasgos ecológicos que disminuyen la detectabilidad en el campo.
En un estudio llevado a cabo en un parque ecológico forestal en Johor, Malasia, Awg Abdul Rahman et al. (2024) investigaron exhaustiva-mente la diversidad de mariposas. Recopilaron datos entre noviembre de 2017 y julio de 2018, utilizando redes aéreas y trampas con cebo a lo largo de un transecto de 1 km. Durante este período, recolectaron 207 individuos pertene-cientes a 75 especies. Encontraron que la familia Nymphalidae fue la más predominante, con 152 individuos distribuidos en 49 especies. Las especies más abundantes fueron Iambrix stellifer Moore, 1878 y Lexias pardalis (Moore, 1878).
Tassin & Goulson (2024) señalan que los jardines son frecuentemente vistos como refugios verdes que ofrecen un santuario para la vida silvestre desplazada del campo por la agricultura intensiva. Su estudio revela que la abundancia y diversidad de mariposas y abejorros en los jardines están significativamente influenciadas por factores tanto extrínsecos como intrínsecos. Además, la gestión adecuada de los jardines puede tener un impacto positivo considerable en las poblaciones de insectos.
Neal et al. (2024) realizaron estudios de transectos lineales y evaluaciones rápidas del hábitat en la estructura del bosque para investigar las asociaciones de hábitat de las mariposas en diez parches de bosque de diferentes tamaños (entre 1 y 40 hectáreas), edades y características en el entorno urbano de Milton Keynes, Reino Unido. Descubrieron que los bosques antiguos albergaban todas las especies detectadas, mientras que las comunidades de mariposas en bosques recreativos y en los bordes de las carreteras mostraban una riqueza, diversidad y abundancia de especies similares, sin importar el tamaño del área. Además, la riqueza de especies de mariposas estaba fuertemente asociada tanto con la superficie del bosque como con la complejidad estructural a escala de sitio, pero solo con la complejidad estructural a escala de transecto. Los resultados sugieren que las técnicas de gestión que aumentan la complejidad estructural dentro de los bosques urbanos pueden ser una manera efectiva de fomentar la riqueza y diversidad de mariposas en contextos donde no es posible aumentar las áreas boscosas.
Para el departamento de Tumbes, Lamas (1976) reportó 121 especies de mariposas diurnas distribuidas en las familias Papilionidae, Pieridae, Nymphalidae, Libytheidae, Lycaenidae, Riodinidae y Hesperiidae; estando representada esta última con 39 especies. Posteriormente se realizaron otras recolecciones registrándose 122 especies en la Zona Reservada (Grados, 1998) y 210 para el departamento (Lamas & Grados, datos inéditos). Después de esos trabajos, no se conoce de la realización de otros estudios sobre diversidad de mariposas en la región.
Por tal razón es que se consideró importante efectuar el presente inventario, llevando a cabo nuevas recolecciones en otras zonas de bosque seco tropical en la Provincia de Zarumilla del Departamento de Tumbes.
MATERIALES Y MÉTODOS
Área de estudio
El Departamento de Tumbes presenta un clima subtropical, correspondiente a una zona de transición entre el ecuatorial y el desértico de la costa peruana. Se considera la región más cálida de la costa peruana, presenta una temperatura promedio casi uniforme en todo el año, siendo las más altas temperaturas promedio las que se presentan de enero a abril (30 °C) y las más bajas o frescas, de junio a septiembre (23 °C). El fenómeno de El Niño y la migración hacia el sur del frente ecuatorial ocasionan la caída de fuertes lluvias estacionales en todo el departamento. Tumbes de acuerdo a las 11 ecorregiones citadas por Brack & Mendiola (2000), se encuentra en la eco-región denominada El Bosque Seco Ecuatorial que comprende una faja costera de 100 a 150 km de ancho en los departamentos de Tumbes, Piura, Lambayeque y La Libertad hasta los 71º Latitud Sur en las vertientes occidentales y la porción seca del valle del Marañón hasta lo 90º LS y en la eco - región denominada Bosque Tropical del Pacífico que se extiende a lo largo de la costa del Pacífico desde el norte del Perú hasta América Central, en el Perú comprende un área poco extensa en el interior del departamento de Tumbes, en El Caucho.
Según la clasificación de zonas de vida de Holdridge (1982), en el territorio tumbesino se hallan las siguientes: Bosque Muy Seco Tropical, Bosque Seco Tropical, Bosque Seco Premontano Tropical, Matorral Desértico Tropical, Monte Espinoso Tropical, y una transicional entre el Matorral Desértico Premontano Tropical a Matorral Desértico Tropical. Los lugares evaluados en el muestreo de mariposas se visitaron desde mayo 2020 hasta abril 2021 en el sector de Matapalo hasta el límite del Bosque de los Manglares de Tumbes, además, de algunos sectores de la margen izquierda del río Zarumilla y de las lagunas Lamederos y Senegal ubicadas en los caseríos de La Coja y Papayal (Tabla 1). Los sectores intervenidos fueron determinados por su cercanía al río Zarumilla que nace del sector Matapalo y desemboca cerca a los Manglares de Puerto 25, además de albergar una vegetación diversa.
Tabla 1
Localidades de muestreo para mariposas, Zarumilla, Tumbes, Perú
|
Sector muestreado |
Lugar |
Elevación (msnm) |
Coordenadas |
|
Matapalo |
Quebrada Faical |
53 |
3° 41' S, 80º 12' W |
|
José Abelardo Quiñones |
Laguna de Quiñones |
34 |
3° 37' S, 80º 12' W |
|
Lechugal |
Orilla del río Zarumilla |
33 |
3° 36' S, 80º 11' W |
|
La Coja |
Laguna Lamederos |
29 |
3° 36' S, 80º 11' W |
|
Papayal |
Laguna Senegal |
24 |
3° 34' S, 80º 13' W |
|
Uña de Gato |
Orilla del río Zarumilla |
21 |
3° 32' S, 80º 13' W |
|
Manglares |
Ingreso a Puerto 25 |
14 |
3° 28' S, 80º 15' W |
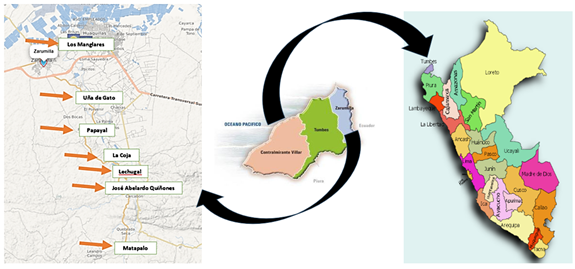
Figura 1. Ubicación de los sectores muestreados en la provincia de Zarumilla durante las evaluaciones de mariposas.
Métodos de muestreo
Instalación de trampas. Las trampas Van Someren-Rydon, fueron instaladas entre 2 a 4 m de altura con respecto al nivel del suelo, según la vegetación presente, y a una distancia de aproximadamente 50 m una de la otra. Se utilizó tres trampas de este tipo. Las trampas fueron instaladas en las zonas muestreadas por un periodo de 24 horas.
Preparación de cebos atrayentes. Se utilizó cebos atrayentes a base de frutas, chicha de jora y melaza o azúcar con proporciones de 70%, 20% y 10% respectivamente. Los cebos fueron colocados en platos descartables en cada trampa Van Someren-Rydon.
Captura de ejemplares con red entomológica. Con ayuda de la red entomológica se complementó la captura de ejemplares de mariposas en los diferentes sitios muestreados, incluyendo aquellos donde se instalaron las trampas. El esfuerzo de muestreo con este método de colecta fue de 40 horas.
Colecta y registro de ejemplares. Para el caso de las trampas Van Someren-Rydon, los especímenes capturados fueron extraídos delicadamente y se les sacrificó haciendo uso de la técnica de la presión dactilar torácica en cada uno de ellos. Algo similar se realizó con los individuos capturados con la red entomológica. Posterior a ello, fueron colocados en sobres de forma triangular de papel milano o de manteca para evitar deteriorar alguna parte de sus cuerpos, como patas, antenas, alas y la pérdida de escamas. Una vez depositado el ejemplar en su sobre, se rotulaba con los datos de recolección: fecha, lugar de captura y nombre del recolector; luego los sobres se guardaron en un recipiente hermético para su posterior traslado y procesamiento.
Identificación. La identificación de la mayoría de las especies la realizó el especialista en mariposas y co-autor del presente trabajo, Dr. Gerardo Lamas, además de contar con el soporte de guías y por comparación con ejemplares ilustrados en la página en línea de las mariposas de América (https://www.butterfliesofamerica.com).
Análisis de datos. Se determinó el número de familias correspondientes a las especies encon-tradas en las diferentes zonas de estudio. La abundancia relativa se determinó según la metodología establecida por Lamas (1985) donde distribuye a las mariposas en cinco clases de abundancia observadas o recolectadas en cierto periodo de muestreo, tal como se muestra en la Tabla 2. Los índices de Shannon-Wiener y de Simpson fueron determinados usando el programa Excel; estos índices son los más utilizados en estudios de ecología ya que combinan la riqueza de especies y su equitatividad (Jost & González-Oreja, 2012).
Tabla 2
Clasificación de abundancia relativa (Lamas 1985)
|
Descripción |
|
|
Especies muy raras (RR) |
Especies muy raras, con menos de seis individuos |
|
Especies raras (R) |
Especies raras, con 6 a 20 individuos. |
|
Especies frecuentes (F) |
Especies frecuentes, con 21 a 50 individuos. |
|
Especies comunes (C) |
Especies comunes, con 51 a 100 individuos. |
|
Especies muy comunes (CC) |
Especies muy comunes, con más de 100 individuos registrados |
RESULTADOS Y DISCUSIÓN
Composición general de la comunidad de mariposas
Se registraron 323 ejemplares de mariposas diurnas, pertenecientes a 69 especies distribuidas en seis familias (Tabla 3) y 17 subfamilias (Tabla 6, Figura 2). Entre las familias que presentaron un mayor número de especies y porcentaje de abundancia (Tabla 3) se encuentran, Nymphalidae (27 especies; 39,13%), Hesperiidae (19 especies; 27,54%) y Pieridae (11 especies; 15,94%). Las subfamilias que presentaron un mayor número de ejemplares por especie y porcentaje de abundancia fueron: Biblidinae(108 ejemplares; 33,44%) y Coliadinae (41 ejemplares; 12,71%). En cuanto a riqueza, las subfamilias con mayor número de especies fueron: Pyrginae y Coliadinae con 10 especies. Las especies Hamadryas amphichloe amphichloe (Hübner, [1821]) (90 ejemplares; 27,9%), Fountainea ryphea ecuadoralis (Johnson & Comstock, 1941) y Anartia amathea sticheli Frushstorfer, 1907 (15 ejemplares; 4,64%) fueron las más abundantes.
En la familia Nymphalidae, la sub familia Biblidinae fue dominante en número de ejemplares de mariposas en la zona de estudio, especialmente por la especie H. amphichloe amphichloe “mariposa tronadora”, (27,86% del total de individuos de dicha subfamilia, Tabla 4). La mayor presencia de esta especie ocurrió en los sectores La Coja, Uña de Gato y Manglares, zonas con una mayor vegetación silvestre arbustiva y poco alteradas por la acción del hombre, en cambio esta especie está ausente en los sectores Quiñones, Lechugal y en el sector Matapalo donde solamente se registró un ejemplar, sectores donde se realiza una mayor actividad antrópica por ser predios agrícolas sembrados de limón y banano donde los productores utilizan herbicidas; además, la deforestación de nuevas áreas para la expansión de la frontera agrícola perjudica a su planta hospedera (denominada “bejuco de fuego”, una Euphorbiaceae del género Dalechampia,) que ocurre en los caminos o cercos de parcelas y de las que se alimentan las larvas de la “mariposa tronadora”. Le siguen la subfamlia Charaxinae con la especie Anaea (Fountainea) ryphea ecuadoralis (4,64%) y Nymphalinae con la especie Anarthia amathea sticheli (4,64%). En la familia Pieridae fue dominante la subfamilia Pierinae con la especie Ascia monuste suasa (Doubleday, 1847) (4,33%), en Hesperiidae, la subfamilia Pyrginae con la especie Heliopetes (Heliopetes) marginata(Edwards, 1882) (2,79%), en Lycaenidae, Theclinae con la especie Strymon davara(Hewitson, 1868) (1,55%), en Riodinidae, la subfamilia Riodinae con la especie Calephelis argyrodines (Bates, 1866) (1,86%) y la Subafmilia con menor dominancia fue Papilioninae con dos especies ambas con 0,93%.
Diversidad de familias
Se identificaron 6 familias de mariposas (Tabla 3) correspondientes a la superfamilia Papilionoidea, entre ellas destacan Nymphalidae (39,13%) y Hesperiidae (27,54). En la Figura 3 se muestran especies representativas de las mariposas por familia, observadas y recolectadas durante el presente estudio. La diversidad de familias de mariposas encontradas en el presente trabajo de investigación es similar a lo reportado por Cárdenas-Lugo et al. (2015) para México en lo que corresponde a número de familias; ellos encontraron cinco familias; en tanto que para el estudio realizado son seis familias, ellos no reportan a la familia Riodinidae, pero hay diferencias en cuanto a los porcentajes por familias. Por otro lado, los resultados son similares en número de familias y porcentajes a lo que reportan Gaviria-Ortiz & Henao-Bañol (2011) para Colombia (Tabla 4).
Lamas (1976) reporta para el departamento de Tumbes una lista de 121 especies de mariposas distribuidas en las familias Papilionidae, Pieridae, Nymphalidae, Lycaenidae, Riodinidae y Hesperiidae siendo similar en cuanto a número de familias al resultado del presente trabajo con excepción de la familia Libytheidae reportada por el citado autor, ahora con rango de subfamilia de Nymphalidae. De esta lista, 17 especies aquí encontradas coinciden con la lista presentada por Lamas (1976) para la provincia de Zarumilla y con 22 especies de la lista reportada por Grados para la Zona Reservada de Tumbes (1988) (Tabla 5). Del resultado del presente trabajo de investigación, con el aporte de 52 especies adicionales, la nueva lista de mariposas teniendo en cuenta el trabajo de Lamas (1976) para Tumbes asciende a 173 especies, aunque según, Lamas & Grados, datos inéditos (Grados, 1998) este número ascendería a 210 para el departamento.
Tabla 3
Familias, riqueza de especies y abundancia de mariposas recolectadas durante el estudio, mayo 2020- abril 2021
|
Nº |
Familias |
# de especies |
Porcentaje (%) |
|
1 |
Nymphalidae |
27 |
39,13 |
|
2 |
Hesperiidae |
19 |
27,54 |
|
3 |
Pieridae |
11 |
15,94 |
|
4 |
Lycaenidae |
7 |
10,14 |
|
5 |
Riodinidae |
3 |
4,35 |
|
6 |
Papilionidae |
2 |
2,90 |
|
Total |
69 |
100 |
|
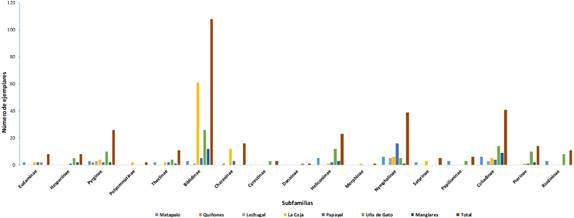
Figura 2. Distribución de mariposas por subfamilias registradas en un área de bosque tropical en la provincia de Zarumilla, departamento de Tumbes, Perú.

Figura 3. Especies representativas de mariposas por familia: (a) Heliconius peruviana C. Felder & R. Felder, 1859 (Nymphalidae); (b) Paches loxus loxus (Westwood, 1852) (Hesperiidae); (c) Hamadryas feronia farinulenta (Fruhstorfer, 1916) (Nymphalidae); (d) Marpesia petreus rheophila Lamas, 1995 (Nymphalidae); (e) Dynamine postverta postverta (Cramer, 1779) (Nymphalidae); (f) Burnsius brenda (Evans, 1942) (Hesperiidae); (g) Pseudolycaena nellyae Lamas, 1981 (Lycaenidae); (h) Proteides mercurius mercurius (Fabricius, 1787) (Hesperiidae).
Tabla 4
Comparación de diversidad de familias y su riqueza con reportes de otros países
|
Cárdenas-Lugo et al. (2015) México |
Gaviria-Ortiz & Henao-Bañol (2011) Colombia |
Estudio realizado Tumbes, Perú |
|
|
Nymphalidae |
53,85% |
45,54% |
38,57% |
|
Pieridae |
32,69% |
10,33% |
15,71% |
|
Papilionidae |
7,69% |
2,82%) |
2,86% |
|
Lycaenidae |
3,85% |
9,86% |
10,00% |
|
Hesperiidae |
1,92% |
23,47% |
30,00% |
|
Riodinidae |
0 |
7,98% |
2,86% |
Tabla 5
Familias y especies de mariposas del presente trabajo de investigación que coinciden con la lista de Lamas (1976) y Grados (1998)
|
Familia |
Especies reportadas por Lamas (1976) |
|
Hesperiidae |
Vehilius limae (Lindsey, 1925) |
|
Gorgythion begga begga, (Prittwitz, 1868) |
|
|
Heliopetes marginata Hayward, 1940 |
|
|
Nisoniades prob. mimas (Cramer, 1775) |
|
|
Paches loxus loxus (Westwood, 1852) |
|
|
Timochares trifasciata trifasciata (Hewitson, 1868) |
|
|
Lycaenidae |
Zizula cyna (Edwards, 1881) |
|
Strymon davara (Hewitson, 1867) |
|
|
Nymphalidae |
Dryas iulia moderata (Riley, 1926) |
|
Heliconius erato cyrbia Godart, 1819 x Heliconius himera Hewitson, 1867 |
|
|
Anartia jatrophae jatrophae (Linnaeus, 1763) |
|
|
Hermeuptychia hermes (Fabricius,1775) |
|
|
Taygetis virgilia (Cramer, 1776) |
|
|
Pieridae |
Eurema elathea ella (Röber, 1909) |
|
Phoebis bourkei (Dixey, 1933) |
|
|
Phoebis marcellina (Cramer, 1777) |
|
|
Ascia monuste suasa (Boisduval, 1836) |
|
|
Familia |
Especies reportadas por Grados (1998) |
|
Hesperiidae |
Gorgythion begga begga, (Prittwitz, 1868) |
|
Paches loxus loxus (Westwood, 1852) |
|
|
Heliopetes arsalte marginata (Hayward, 1940) |
|
|
Lycaenidae |
Pseudolycaena nellyae (Lamas, 1981) |
|
Nymphalidae |
Dryas iulia moderata (Riley, 1926) |
|
Heliconius erato cyrbia Godart, 1819 x Heliconius himera Hewitson, 1867 |
|
|
Dione juno andicola (Bates, 1864) |
|
|
Anartia amathea sticheli (Fruhstorfer, 1907) |
|
|
Anartia jatrophae jatrophae (Linnaeus, 1763) |
|
|
Junonia genoveva infuscata (C & Felder, 1861) |
|
|
Hamadryas amphichloe amphichloe (Boisduval, 1870) |
|
|
Mestra dorcas latimargo (Hall, 1929) |
|
|
Dynamine postverta postverta (Cramer, [1979]) |
|
|
Marpesia petreus reophila (Lamas, 1995) |
|
|
Fountainea ryphea ecuadoralis (Johnson & Comstock, 1941) |
|
|
Hermeuprychia hermes (Fabricius, 1775) |
|
|
Danaus eresimus erginus (Godman & Salvin, 1897) |
|
|
Pieridae |
Pyrisitia nise stygma (Boisduval, 1836) |
|
Pyrisitia proterpia (Fabricius, 1775) |
|
|
Riodinidae |
Calephelis argyrodine (Bates, 1856) |
|
Papilionidae |
Heraclides anchisiadis lamasi (Brown, 1994) |
|
Heraclides paeon thrason (C & Felder, 1865) |
|
|
Proteidus mercurius mercurius (Fabricius, 1787) |
Riqueza de especies
La riqueza de mariposas recolectadas e identificadas en la presente investigación fue de 69 especies (Tabla 6), 37 están identificadas a nivel de subespecie, 29 a nivel de especie, una a nivel de género, una a nivel de familia y una corresponde a un hibrido inter-específico. En esta tabla se presenta una lista detallada de mariposas. En la familia Hesperiidae se encontraron 19 especies distribuidas entre las subfamilias Eudaminae, Hesperiinae y Pyrginae. La familia Lycaenidae cuenta con siete especies distribuidas en las subfamilias Polyommatinae y Theclinae. En cuanto a la familia Nymphalidae, se identificaron 27 especies distribuidas en las subfamilias Biblidinae, Charaxinae, Cyrestinae, Danainae, Heliconiinae, Morphinae, Nymphalinae y Satyrinae. La familia Papilionidae incluye dos especies de la subfamilia Papilioninae, mientras que la familia Pieridae comprende 11 especies distribuidas en las subfamilias Coliadinae y Pierinae. Finalmente, en la familia Riodinidae se encontraron tres especies de la subfamilia Riodininae.
El conocimiento de la riqueza de especies de mariposas en un determinado hábitat es impor-tante para evaluar impactos ambientales, así, por ejemplo; Kaziske et al. (2023) utilizando datos nacionales del Programa Alemán de Monitoreo de Mariposas, analizaron el impacto de tres indica-dores relacionados con la cobertura del suelo y la intensidad del uso agrícola en la riqueza de especies y en la composición de rasgos de las comunidades de mariposas. Los resultados coinci-dieron con estudios previos en áreas geográficas más pequeñas, subrayando la importancia de mantener una baja densidad de ganado herbívoro para evitar la pérdida de insectos polinizadores y preservar tanto la biodiversidad como los servicios ecosistémicos en los paisajes agrícolas.
Otro aspecto a tener en cuenta es el clima, por ejemplo, las expectativas basadas en el clima para las próximas décadas son negativas para un número importante de mariposas europeas (Lepidoptera, superfamilia Papilionoidea) (Settele et al. 2008; Warren et al., 2021).
Por otro lado, Fumy & Fartmann (2023) analizaron la riqueza de especies de mariposas y saltamontes amenazados en ecosistemas de turberas y pastizales en una cadena montañosa baja en el suroeste de Alemania, específicamente en el sur de la Selva Negra. Investigaron un total de 84 parcelas seleccionadas al azar (100 m x 100 m). Cada parcela, situada a lo largo de un gradiente hidrológico, correspondía a uno de los cinco tipos de hábitat: turbera, pantano, pastizal mésico, pastizal semiseco y pastizal seco. Ellos identificaron diferencias significativas en las condiciones ambientales y en la composición de especies amenazadas de mariposas y saltamontes entre hábitats de turberas y pastizales. La riqueza de especies y el número de especies indicadoras alcanzaron su punto máximo en pantanos y pastizales secos, y en menor medida en pastizales semisecos. Estos tres tipos de hábitat se caracte-rizaron por niveles de uso de la tierra de bajos a intermedios. Consistentemente, la intensidad del uso de la tierra resultó ser el factor clave que determinó la heterogeneidad del hábitat y, en consecuencia, la riqueza de especies de mariposas y saltamontes amenazados.
Abundancia relativa (Fa)
En la Tabla 6 se muestra la abundancia relativa de cada una de las especies recolectadas en cada uno de los sectores evaluados, siendo los sectores Uña de gato (102) y La Coja (101) donde se recolectó el mayor número de ejemplares y el menor número en el sector Quiñones (3). Con respecto a la abundancia relativa se registró 57 especies que corresponden a la clase de “muy raras” (RR), 12 especies a “raras” (R) y una especie que representa la clase “común” (C). La familia Hesperiidae tuvo mayor abundancia (13,02%), siendo los sectores con mayor número de especies Uña de Gato y La Coja y el sector de Matapalo. En cuanto al número de individuos, los sectores de Uña de Gato y La Coja fueron los más representativos. En cambio, la familia con menos abundancia fue Papilionidae con 1,86% con un número de especies e individuos muy bajo en el sector Quiñones. Al respecto, cabe indicar que esta familia es un grupo de mariposas con alto valor ecológico y de mayor interés para la conservación a nivel mundial (Yu et al., 2023). La especie más común y atraída al cebo cuando se instalaron las trampas Van Someren-Rydon, en los distintos sectores evaluados, fue H. amphichloe amphichloe (Nymphalidae: Biblidinae) conocida comúnmente como “mariposa tronadora.
En Papilionidae, las especies Heraclides paeon thrason (Felder & Felder, 1865) y H. anchisiades lamasi (Brown, 1994) se consideran muy raras por presentar una abundancia de 1,86%. En Pieridae, la especie A. monuste suasa con 4,33% se clasifica como una especie rara. En Riodinidae, las dos únicas especies fueron Calephelis argyrodines y Lasaia agesilas agesilas (Latreille, [1833]) con porcentajes de abundancia de 1,86% y 1,24% respectivamente. La primera fue considerada rara, y muy rara la segunda.
De acuerdo a sus preferencias ecológicas princi-pales, las mariposas pueden ser divididas en dos grandes grupos: (i) mariposas pertenecientes a formaciones vegetales “abiertas” (preponderancia de especies de Pieridae, Lycaenidae y Hesperiidae), y (ii) mariposas de formaciones vegetales “cerradas” (predominancia de Nymphalidae) (Lamas, 1976), el mismo autor indica que formaciones “abiertas” son aquellas en las cuales la luz del suelo llega al suelo en gran cantidad, y “cerradas” aquellas en las que un “dosel” formado por las copas adyacentes árboles impide que la mayor parte de la radiación solar llegue directamente al suelo. En general, los hábitats de los sectores La Coja y Uña de Gato resultaron ser de mayor importancia para la conservación de la riqueza y diversidad de especies de mariposas, esto probablemente se debe a que representan el hábitat original de la zona, que mantiene una alta riqueza, diversidad y estructura florística, situación que crea condiciones micro climáticas ideales para adultos de mariposas de áreas cerradas que no toleran la luz solar de manera directa (DeVries, 1987). Con relación a esto, en este trabajo la mayor parte de especies recolectadas pertenecen a la familia Nymphalidae y en los sectores La Coja (89), Uña de Gato (47), Papayal (26), Manglares y Matapalo (16), es decir, son mariposas que prefieren las formaciones vegetales cerradas. En el caso de las mariposas que prefieren las formaciones vegetales “abiertas”, las familias más importantes han sido Hesperiidae (19) y Pieridae (11) recolectadas en el sector Uña de Gato.
Con relación a la ocurrencia de H. amphichloe ampichloe, esta especie es citada por Brito & Buestán (2014) para el Bosque Protector Cerro Blanco (BPCB), un bosque seco de la costa ecuatoriana, indicando que presenta juntamente con la especie H. amphinome (Linnaeus, 1767) alta abundancia siendo ambas relativamente sencillas de recolectar e identificar en el campo. A. monuste suasa es referida por Gareca & Reichle (2010) para Bolivia; en tanto que C. argyrodines es reportada por Salinas-Gutiérrez et al. (2009) en un listado de Papilionoidea para Guatemala.
Respecto a la especie Strymon davara, Peña & Ugarte (1996) indican que es muy poco conocida con una distribución restringida al Sur de Perú y Norte de Chile. Robbins & Nicolay (2002) a su vez expresan que, como también es observado en muchas otras especies del género, los machos de esta especie ocupan territorios de apareamiento donde se posan. Vargas & Duarte (2016) en estudios realizados en el desierto de Atacama, específicamente en el valle de Azapa, provincia de Arica, al norte de Chile, reportan haber encontrado durante los años 2002 y 2015 larvas de esta especie alimentándose de inflorescencias de la maleza Alternanthera halimifolia (Lam.) Standl., 1931 (Amaranthaceae). Consideran como un ejemplo la importancia de esta planta en la conservación de las poblaciones locales de una mariposa en un entorno altamente modificado por el hombre.
Ma et al. (2015) manifiestan que las malezas pueden estar involucradas en muchas interac-ciones en los agroecosistemas. La competencia con los cultivos es quizás lo más evidente, debido a su impacto económico. Sin embargo, las malezas proveen servicios extremadamente importantes, como fuente de alimento o refugio para polini-zadores (Nichols & Altieri, 2013) entre los cuales se encuentran las mariposas. Con relación a las especies H. anchisiades lamasi y H. paeon thrason, Vásquez et al. (2017) en un estudio realizado sobre plantas alimenticias para mariposas, destacan que estas especies son comunes en áreas deforestadas. La primera de ellas está asociada a la planta Citrus medica L., 1753 (Rutaceae) y la segunda a Piper aduncum L., 1753 (Piperaceae).
Se apreció que en los sectores evaluados existen diferencias en la estructura y composición florística, lo que se refleja en las diferencias observadas en la diversidad y composición de mariposas. Sin duda alguna, el factor antrópico es causante de gran importancia por su intervención en la modificación y acondicionamiento de los bosques para fines propios, afectando a las comunidades de mariposas y, posiblemente su desplazamiento a zonas más altas de las montañas haciendo que ciertas especies solo puedan ser vistas en dichas áreas. Kurylo et al., (2020) en un estudio realizado en Melbourne Australia, encon-traron que el incremento en la cobertura arbórea contribuyó a un mayor número de especies de mariposas y que los efectos adversos que provoca la creciente urbanización en la riqueza y abundancia de mariposas pueden mitigarse, en cierta medida, mejorando la calidad del entorno urbano; así mismo destacan que esto puede lograrse incrementando la cantidad de árboles y proporcionando recursos florales. Además, resaltan que hay pruebas que indican que las plantas autóctonas o nativas resultan ser más efectivas para este propósito.
Esfuerzos futuros de conservación deberían centrarse en especies con contracción de hábitat, distribución limitada y endemicidad con medidas tanto in situ como ex situ, especialmente en áreas protegidas (Yu et al., 2023).
Tabla 6
Riqueza y abundancia relativa de especies de mariposas recolectadas en los sectores de Matapalo, Quiñones, Lechugal, La Coja, Papayal, Uña de Gato y Los Manglares desde mayo de 2020 a abril de 2021 en la provincia de Zarumilla
|
Nº |
Familia /subfamilia/especie |
Sectores evaluados |
Total, por especie |
% de abundancia |
Clasificación |
||||||
|
Matapalo |
Quiñonez |
Lechugal |
La Coja |
Papayal |
Uña de gato |
Manglares |
|||||
|
|
Hesperiidae |
|
|
|
|
|
|
|
|
|
|
|
|
Eudaminae |
|
|
|
|
|
|
|
|
|
|
|
1 |
Cecropterus (Thorybes) dorantes dorantes (Stoll, 1790) |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
2 |
Chioides catillus jethira (Butler, 1870) |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
3 |
Codatractus (Caudatractus) apulia Evans, 1952 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
4 |
Proteides mercurius mercurius (Fabricius, 1787) |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
5 |
Spicauda procne (Plötz, 1881) |
0 |
0 |
0 |
2 |
2 |
0 |
0 |
4 |
1,24 |
RR |
|
|
Sub total por subfamilia y sector |
2 |
0 |
0 |
2 |
2 |
2 |
0 |
8 |
2,48 |
|
|
|
Hesperiinae |
|
|
|
|
|
|
|
|
|
|
|
6 |
Cyamenes limae (Lindsey, 1925) |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
0,31 |
RR |
|
7 |
Hedone vibex praeceps Scudder, 1872 |
0 |
0 |
0 |
0 |
0 |
2 |
1 |
3 |
0,93 |
RR |
|
8 |
Niconiades nikko Hayward, 1948 |
0 |
0 |
0 |
0 |
0 |
2 |
0 |
2 |
0,62 |
RR |
|
9 |
Pompeius pompeius (Latreille, 1824) |
0 |
0 |
0 |
0 |
1 |
0 |
0 |
1 |
0,31 |
RR |
|
10 |
ejemplar no ID |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
|
Sub total por subfamilia y sector |
0 |
0 |
0 |
0 |
1 |
5 |
2 |
8 |
2,48 |
|
|
|
Pyrginae |
|
|
|
|
|
|
|
|
|
|
|
11 |
Burnsius brenda (Evans, 1942) |
0 |
0 |
1 |
0 |
1 |
2 |
0 |
4 |
1,24 |
RR |
|
12 |
Eantis thraso (Hübner, 1807) |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
2 |
0,62 |
RR |
|
13 |
Gorgythion begga begga (Prittwitz, 1868) |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
14 |
Heliopetes marginata Hayward, 1940 |
2 |
0 |
1 |
3 |
0 |
3 |
0 |
9 |
2,79 |
R |
|
15 |
Nisoniades prob. mimas (Cramer, 1775) |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
16 |
Nisoniades rubescens (Moschler, 1877) |
0 |
0 |
1 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
17 |
Paches loxus loxus (Westwood, 1852) |
0 |
2 |
0 |
0 |
0 |
0 |
0 |
2 |
0,62 |
RR |
|
18 |
Timochares trifasciata trifasciata (Hewitson, 1868) |
0 |
0 |
0 |
0 |
0 |
3 |
1 |
4 |
1,24 |
RR |
|
19 |
Xenophanes tryxus (Stoll, 1780) |
1 |
0 |
0 |
0 |
1 |
0 |
0 |
2 |
0,62 |
RR |
|
|
Subtotal por subfamilia y sector |
3 |
2 |
3 |
4 |
2 |
10 |
2 |
26 |
8,06 |
|
|
|
Total, por familia y sector |
5 |
2 |
3 |
6 |
5 |
17 |
4 |
42 |
13,02 |
|
|
|
Lycaenidae |
|
|
|
|
|
|
|
|
|
|
|
|
Polyommatinae |
|
|
|
|
|
|
|
|
|
|
|
20 |
Zizula cyna (Edwards, 1881) |
0 |
0 |
0 |
2 |
0 |
0 |
0 |
2 |
0,62 |
RR |
|
|
Subtotal por subfamilia y sector |
0 |
0 |
0 |
2 |
0 |
0 |
0 |
2 |
0,62 |
|
|
|
Theclinae |
|
|
|
|
|
|
|
|
|
|
|
21 |
Arawacus lincoides (Draudt, 1917) |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
22 |
Calycopis bactra (Hewitson, 1877) |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,32 |
RR |
|
23 |
Calycopis lerbela Field, 1967 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,33 |
RR |
|
24 |
Pseudolycaena nellyae Lamas, 1981 |
0 |
0 |
0 |
0 |
0 |
2 |
0 |
2 |
0,62 |
RR |
|
25 |
Siderus parvinotus Kaye, 1904 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
26 |
Strymon davara (Hewitson, 1867) |
0 |
0 |
0 |
2 |
2 |
0 |
1 |
5 |
1,55 |
RR |
|
|
Subtotal por subfamilia y sector |
2 |
0 |
0 |
2 |
2 |
4 |
1 |
11 |
3,41 |
|
|
|
Total, por familia y sector |
2 |
0 |
0 |
4 |
2 |
4 |
1 |
13 |
4,03 |
|
|
|
Nymphalidae |
|
|
|
|
|
|
|
|
|
|
|
|
Biblidinae |
|
|
|
|
|
|
|
|
|
|
|
27 |
Dynamine haenschi Hall, 1917 |
0 |
0 |
0 |
0 |
1 |
0 |
0 |
1 |
0,31 |
RR |
|
28 |
Dynamine postverta postverta (Cramer, 1779) |
2 |
0 |
0 |
0 |
0 |
1 |
0 |
3 |
0,93 |
RR |
|
29 |
Hamadryas amphichloe amphichloe (Boisduval, 1870) |
1 |
0 |
0 |
54 |
2 |
21 |
12 |
90 |
27,86 |
C |
|
30 |
Hamadryas amphinome fumosa (Fruhstorfer, 2015) |
0 |
0 |
0 |
1 |
0 |
1 |
0 |
2 |
0,62 |
RR |
|
31 |
Hamadryas feronia farinulenta (Fruhstorfer, 1916) |
0 |
0 |
0 |
6 |
2 |
0 |
0 |
8 |
2,48 |
R |
|
32 |
Mestra dorcas latimargo (Hall, 1929) |
0 |
0 |
1 |
0 |
0 |
3 |
0 |
4 |
1,24 |
RR |
|
|
Subtotal por subfamilia y sector |
3 |
0 |
1 |
61 |
5 |
26 |
12 |
108 |
33,44 |
|
|
|
Charaxinae |
|
|
|
|
|
|
|
|
|
|
|
33 |
Archaeoprepona amphimachus amphiktion (Fruhstorfer, 1916) |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
34 |
Fountainea ryphea ecuadoralis (Johnson & Comstock, 1941) |
0 |
1 |
0 |
11 |
3 |
0 |
0 |
15 |
4,64 |
R |
|
|
Subtotal por subfamilia y sector |
0 |
1 |
0 |
12 |
3 |
0 |
0 |
16 |
4,95 |
|
|
|
Cyrestinae |
|
|
|
|
|
|
|
|
|
|
|
35 |
Marpesia petreus rheophila Lamas, 1995 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0,93 |
RR |
|
|
Subtotal por subfamilia y sector |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0,93 |
|
|
|
Danainae |
|
|
|
|
|
|
|
|
|
|
|
36 |
Danaus eresimus erginus (Godman & Salvin, 1897) |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
|
Subtotal por subfamilia y sector |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
|
|
|
Heliconiinae |
|
|
|
|
|
|
|
|
|
|
|
37 |
Dione juno andicola (Bates, 1864) |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0,93 |
RR |
|
38 |
Dione vanillae forbesi (Michener, 1942) |
0 |
0 |
0 |
0 |
1 |
3 |
3 |
7 |
2,17 |
R |
|
39 |
Dryas iulia moderata (Riley, 1926) |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
40 |
Eueides isabella ecuadorensis Strand, 1912 |
1 |
0 |
0 |
0 |
0 |
1 |
0 |
2 |
0,62 |
RR |
|
41 |
Euptoieta hegesia meridiania Stichel, 1938 |
0 |
0 |
0 |
1 |
0 |
1 |
0 |
2 |
0,62 |
RR |
|
42 |
Heliconius erato cyrbia Godart, 1819 x Heliconius himera Hewitson, 1867 |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
43 |
Heliconius peruviana C. Felder & R. Felder, 1859 |
4 |
0 |
0 |
0 |
1 |
2 |
0 |
7 |
2,17 |
R |
|
|
Subtotal por subfamilia y sector |
5 |
0 |
0 |
1 |
2 |
12 |
3 |
23 |
7,13 |
|
|
|
Morphinae |
|
|
|
|
|
|
|
|
|
|
|
44 |
Morpho helenor cormieri Blandin, 2007 |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
|
Subtotal por subfamilia y sector |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
1 |
0,31 |
|
|
|
Nymphalinae |
|
|
|
|
|
|
|
|
|
|
|
45 |
Anartia amathea sticheli Fruhstorfer, 1907 |
4 |
0 |
4 |
1 |
6 |
0 |
0 |
15 |
4,64 |
R |
|
46 |
Anartia jatrophae jatrophae (Linnaeus, 1763) |
0 |
0 |
0 |
2 |
3 |
0 |
0 |
5 |
1,55 |
RR |
|
47 |
Anthanassa frisia taeniata (Röber, 1913) |
1 |
0 |
0 |
1 |
1 |
4 |
0 |
7 |
2,17 |
R |
|
48 |
Eresia eunice drypetis Godman & Salvin, 1878 |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
49 |
Junonia genoveva infuscata C. Felder & R. Felder, 1867 |
0 |
0 |
1 |
2 |
6 |
1 |
1 |
11 |
3,41 |
R |
|
|
Subtotal por subfamilia y sector |
6 |
0 |
5 |
6 |
16 |
5 |
1 |
39 |
12,08 |
|
|
|
Satyrinae |
|
|
|
|
|
|
|
|
|
|
|
50 |
Hermeuptychia hermes (Fabricius,1775) |
0 |
0 |
0 |
1 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
51 |
Malaveria sp. |
0 |
0 |
0 |
2 |
0 |
0 |
0 |
2 |
0,62 |
RR |
|
52 |
Taygetis virgilia (Cramer, 1776) |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
53 |
Yphthimoides renata (Stoll, 1780) |
1 |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
0,31 |
RR |
|
|
Subtotal por subfamilia y sector |
2 |
0 |
0 |
3 |
0 |
0 |
0 |
5 |
1,55 |
|
|
|
Total, por familia y sector |
16 |
1 |
6 |
84 |
26 |
47 |
16 |
196 |
60,7 |
|
|
|
Papilionidae |
|
|
|
|
|
|
|
|
|
|
|
|
Papilioninae |
|
|
|
|
|
|
|
|
|
|
|
54 |
Heraclides anchisiades lamasi Brown, 1994 |
0 |
0 |
0 |
0 |
0 |
3 |
0 |
3 |
0,93 |
RR |
|
55 |
Heraclides paeon thrason (C. Felder & R. Felder, 1865) |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0,93 |
RR |
|
|
Subtotal por familia y sector |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
6 |
1,86 |
|
|
|
Total, por familia y sector |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
6 |
1,86 |
|
|
|
Pieridae |
|
|
|
|
|
|
|
|
|
|
|
|
Coliadinae |
|
|
|
|
|
|
|
|
|
|
|
56 |
Abaeis (Lucidia) albula totora (Lamas, 1981) |
0 |
0 |
2 |
0 |
0 |
0 |
0 |
2 |
0,62 |
RR |
|
57 |
Abaeis (Sphaenogona) arbela angulata (Wallengren, 1860) |
0 |
0 |
0 |
3 |
0 |
0 |
0 |
3 |
0,93 |
RR |
|
58 |
Anteos maerula (Fabricius, 1775) |
0 |
0 |
0 |
0 |
0 |
4 |
0 |
4 |
1,24 |
RR |
|
59 |
Eurema elathea ella (Röber, 1909) |
1 |
0 |
0 |
0 |
3 |
4 |
3 |
11 |
3,41 |
R |
|
60 |
Phoebis agarithe tumbesina Lamas, 1981 |
0 |
0 |
0 |
2 |
0 |
2 |
1 |
5 |
1,55 |
RR |
|
61 |
Phoebis argante larra (Fabricius, 1798) |
0 |
0 |
0 |
0 |
0 |
2 |
0 |
2 |
0,62 |
RR |
|
62 |
Phoebis bourkei (Dixey, 1933) |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
0,31 |
RR |
|
63 |
Phoebis marcellina (Cramer, 1777) |
1 |
0 |
0 |
0 |
0 |
0 |
1 |
2 |
0,62 |
RR |
|
64 |
Pyrisitia nise stygma (Boisduval, 1836) |
1 |
0 |
1 |
0 |
1 |
2 |
3 |
8 |
2,48 |
R |
|
65 |
Pyrisitia proterpia (Fabricius, 1775) |
3 |
0 |
0 |
0 |
0 |
0 |
0 |
3 |
0,93 |
RR |
|
|
Subtotal por subfamilia y sector |
6 |
0 |
3 |
5 |
4 |
14 |
9 |
41 |
12,71 |
|
|
|
Pierinae |
|
|
|
|
|
|
|
|
|
|
|
66 |
Ascia monuste suasa (Boisduval, 1836) |
0 |
0 |
0 |
1 |
1 |
10 |
2 |
14 |
4,33 |
R |
|
|
Subtotal por subfamilia y sector |
0 |
0 |
0 |
1 |
1 |
10 |
2 |
14 |
4,33 |
|
|
|
Total, por familia y sector |
6 |
0 |
3 |
6 |
5 |
24 |
11 |
55 |
17,04 |
|
|
|
Riodinidae |
|
|
|
|
|
|
|
|
|
|
|
|
Riodininae |
|
|
|
|
|
|
|
|
|
|
|
67 |
Calephelis argyrodines (Bates, 1866) |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
6 |
1,86 |
R |
|
68 |
Lasaia agesilas agesilas (Latreille, 1809) |
0 |
0 |
0 |
0 |
0 |
4 |
0 |
4 |
1,24 |
RR |
|
69 |
Melanis leucophlegma (Stichel, 1910) |
0 |
0 |
0 |
0 |
0 |
1 |
0 |
1 |
0,31 |
RR |
|
|
Subtotal por subfamilia y sector |
3 |
0 |
0 |
0 |
0 |
8 |
0 |
11 |
3,41 |
|
|
|
Total, por familia y sector |
3 |
0 |
0 |
0 |
0 |
8 |
0 |
11 |
3,41 |
|
|
|
Total, por Sector evaluado |
35 |
3 |
12 |
100 |
38 |
103 |
32 |
323 |
100,00 |
|
Índices de diversidad de Shannon (H) y de Simpson (ʎ)
En la Tabla 7 se muestran los índices de diversidad de Shannon y Simpson. Según el índice de Shannon, los valores que se obtuvieron en los sectores de Uña de Gato con 3,21; en Matapalo con 2,88; en Papayal 2,61; en Los Manglares con 2,18; en La Coja con 1,95; en Lechugal con 1,91 y en Quiñones con 0,64. Esto indica que, con valores superiores a 3, la diversidad de especies correspondientes al sector Uña de Gato es alta. En el caso de los sectores de Matapalo, Papayal y Los Manglares sus valores se encuentran en el rango de 2 y 3 indicando que la diversidad se encuentra en equilibrio. Finalmente, los sectores de La Coja, Lechugal y Quiñones exhiben valores menores a 2, indicando que la diversidad de mariposas es baja. Para el índice Simpson, los mayores valores cercanos a 1 corresponden a los sectores de Matapalo, Uña de Gato, Papayal, Lechugal, Manglares y La Coja con 0,93; 0,93; 0,91; 0,82; 0,82 y 0,69 respectivamente; indicando que son sectores con mayor diversidad de especies y que la población de una especie de mariposa diurna predomine sobre las demás; sin embargo, en el sector de Quiñones se obtuvo un valor de 0,44 cercano a 0, indicando un sector con menor diversidad y una dominancia relativamente regular de algunas especies de mariposas.
Comprender los factores que influyen en los patrones de diversidad, riqueza y densidad de los organismos es fundamental para gestionar la biodiversidad en el contexto del cambio global (Barea-Azcón et al., 2023).
Tabla 7
Índices de diversidad de mariposas por familia recolectadas en los sectores Matapalo, Quiñones, Lechugal, La Coja, Papayal, Uña de Gato y Los Manglares desde mayo del 2020 a abril de 2021 en la provincia de Zarumilla
|
Familias |
Sectores de evaluación |
Total |
||||||
|
Matapalo |
Quiñonez |
Lechugal |
La Coja |
Papayal |
Uña de Gato |
Manglares |
||
|
Nº de individuos por familia por sector |
||||||||
|
Nymphalidae |
16 |
1 |
6 |
84 |
26 |
47 |
16 |
196 |
|
Pieridae |
6 |
0 |
3 |
6 |
5 |
24 |
11 |
55 |
|
Hesperiidae |
5 |
2 |
3 |
7 |
4 |
17 |
4 |
42 |
|
Lycaenidae |
2 |
0 |
0 |
4 |
2 |
4 |
1 |
13 |
|
Riodinidae |
3 |
0 |
0 |
0 |
0 |
8 |
0 |
11 |
|
Papilionidae |
3 |
0 |
0 |
0 |
0 |
3 |
0 |
6 |
|
Número total de individuos por sector |
35 |
3 |
12 |
101 |
37 |
103 |
32 |
323 |
|
Número de especies |
21 |
2 |
8 |
22 |
18 |
37 |
14 |
|
|
Diversidad especifica |
5,63 |
0,91 |
2,82 |
4,55 |
4,71 |
7,77 |
3,75 |
|
|
Índice de Shanoon (H) |
2,88 |
0,64 |
1,91 |
1,95 |
2,61 |
3,21 |
2,18 |
|
|
Índice de Simpson (ʎ) |
0,93 |
0,44 |
0,82 |
0,69 |
0,91 |
0,93 |
0,82 |
|
CONCLUSIONES
En el presente trabajo de investigación se logró encontrar seis familias de mariposas diurnas de la superfamilia Papilionoidea que son: Nymphalidae (38,57%), Hesperiidae (30,00%), Pieridae (15,71%), Lycaenidae (10,00%), Papilionidae (2,86%) y Riodinidae (2,86%); siendo la primera familia con más riquezas de especies y abundancia para un área de bosque tropical.
Durante el proceso de investigación, se encontró un hibrido interespecífico de mariposa. Sin embargo, se presume que aún existen muchas especies de mariposas por identificar en la parte alta del bosque, creyéndose que posiblemente sean nuevos registros para la región Tumbes. Asimismo, se recomienda realizar investigaciones con respecto a las plantas hospederas de las mariposas e implementar un manual de identificación de estas.
REFERENCIAS BIBLIOGRÁFICAS
Alanen, E., Hyvönen, T., Lindgren, S., Härmä, O., & Kuussaari, M. (2011). Differential responses of bumblebees and diurnal Lepidoptera to vegetation succession in long-term set-aside. J Appl Ecol 48(5):1251–1259.
Amat-García, G., Andrade-Correa, M. G., & Amat-García, E. (2007). Libro Rojo de los invertebrados terrestres de Colombia. Bogotá: Universidad Nacional de Colombia. 215 pp.
Andrade-Correa, M. G. (1998). Utilización de las mariposas como bioindicadoras del tipo de hábitat y su biodiversidad. Colombia. Revista de la Academia Colombiana de Ciencias Exactas, Físicas y Naturales, 22(84), 407-421.
Awg Abdul Rahman, A., Ahmad Fahmi, U. S., Suhaili, K., & Mohamed, M. (2024). Butterflies species richness and diversity in Panti Forest Eco-Park, Johor, Malaysia. BIO web of conferences, 94, 04002. https://doi.org/10.1051/bioconf/20249404002.
Barea-Azcón, J. M., Pérez-Luque, A. J., Olivares, F. J., Guerrero, M., Galiana-García, M., Chaves, M. D., Olvera, M., & Munguira, M. L. (2023). Butterfly diversity, richness, and density patterns in Sierra Nevada (SE Spain): Conservation implications under a global change scenario. Insect Conservation and Diversity, 16(2), 217–230. https://doi.org/10.1111/icad.12617.
Blair, R. B. (1999). Birds and butterflies along an urban gradient: Surrogate taxa for assessing biodiversity? Ecological Applications, 9, 164–170. https://doi.org/10.2307/2641176
Boggs, C. L., Watt, W. B., & Ehrlich, P. R. (2003). Butterflies: ecology and evolution taking flight. The University of Chicago Press, Chicago.
Bonebrake, T., Ponisio, L., Boggs, C., & Ehrlich, P. (2010). More than just indicators: A review of tropical butterfly ecology and conservation. Biological Conservation, 143, 1831-1841.
Brack, A., & Mendiola, C. (2000). Ecología del Perú. Parte II. Las regiones naturales del Perú. Lima, Editorial Bruño. 169 pp.
Brito, G., & Buestán, J. (2014). Diversidad y distribución de lepidópteros diurnos ropaloceros en cinco categorías de vegetación y dos estratos de bosque (Sotobosque - subdosel) en el bosque protector Cerro Blanco Guayquil - Ecuador. Guayaquil, Ecuador. Revista Científica Ciencias Naturales y Ambientales, 8(1), 9-16.
Brown, K. (1997). Diversity, disturbance, and sustainable use of Neotropical forests: insects as indicators for conservation moni-toring. Journal of Insect Conservation, 1, 25-42.
Brown, K. S., & Freitas, A. V. L. (2000). Atlantic forest butterflies: indicators for landscape conservation. Biotropica, 32(4b), 934-956.
Cárdenas-Lugo, C. P., León-Cortés, J. L., & Angulo-Audeves, J. T. (2015). Diversidad, distribución y abundancia de mariposas en hábitats costeros de Sinaloa, México (Insecta: Lepidoptera) SHILAP Revista de Lepidopterología, 43(169), 15-26.
Comay, O., Ben Yehuda, O., Schwartz-Tzachor, R., Benyamini, D., Pe’er, I., Ktalav, I., & Pe’er, G. (2021). Environmental controls on butterfly occurrence and species richness in Israel: The importance of temperature over rainfall. Ecology and Evolution, 11(17), 12035–12050. https://doi.org/10.1002/ece3.7969
DeVries, P. J. (1987). The Butterflies of Costa Rica and Their Natural History: Papilionidae, Pieridae, Nymphalidae. Princeton, Princeton University Press. 327 pp.
DeVries, P. J., Walla, T. R., & Greeney, H. (1997). Species diversity in spatial and temporal dimensions of fruit feeding butterflies from two Ecuadorian rainforests. Biol. J. Linn. Soc., 68, 333-353.
DeVries, P. J., Walla, T. R., & Greeney, H. (1999). Species diversity in spatial and temporal dimensions of fruit feeding butterflies from two Ecuadorian rainforests. Biol. J. Linn. Soc., 68, 333-353.
DeVries, P. J., & Walla, T.R. (2001). Species diversity and community structure in Neotropical fruit-feeding butterflies. Biological Journal of the Linnean Society, 74(1), 1-15.
Fumy, F., & Fartmann, T. (2023). Low-intensity land use fosters species richness of threatened butterflies and grasshoppers in mires and grasslands. Global Ecology and Conservation, 41, e02357. https://doi.org/10.1016/j.gecco.2022.e02357.
Garrido, P., Naumov, V., Söderquist, L., et al. (2022). Effects of experimental rewilding on butterflies, bumblebees and grasshoppers. J Insect Conserv, 26, 763–771 https://doi.org/10.1007/s10841-022-00420-4
Gasca, J., Tovar, M., Ávila - Cubillos, C., Viveros -Mafla, J., & Beltrán-Solano, Y. A. ( 2015). Análisis de la microestructura de dos especies de lepidópteras para modelos de prototipo en la construcción. Agroecol. Cienc. Tecnol., 3 (1), 7-12.
Gaviria-Ortiz, F. G., & Henao-Bañol, E. R. (2011). Diversidad de mariposas diurnas (Hesperioidea-Papilionoidea) del Parque Natural Regional el vínculo (Buga-Vale del Cauca). Boletín Científico. Centro de Museos. Museo de Historia Natural, 15(1), 115-133.
Gilbert, L. E. (1972). Pollen feeding and reproductive biology of Heliconius butterflies. Proceedings of the National Academy of Sciences of the United States of America 69 (6): 1403-1407.
Goulson, D., Hanley, M. E., Darvill, B., Ellis, J., & Knight, M. E. (2005). Causes of rarity in bumblebees. Biol Conserv, 122(1), 1–8.
Grados. J. (1998). Informe del Inventario de la fauna de mariposas del Bosque nacional de Tumbes pp. 89-93. In Wust W. (Ed). Zona Reservada de Tumbes, Biodiversidad y Diagnóstico Socioeconómico. Lima, PROFONANPE).
Holdridge, L. (1982). Ecología basada en zonas de vida. San José, IICA. 216 pp.
Jost, L., & González-Oreja, J. (2012). Midiendo la diversidad biológica: más allá del índice de Shannon. Acta Zoológica Lilloana, 56(1-2), 3–14.
Kasiske, T., Dauber, J., Harpke, A., Klimek, S., Kühn, E., Settele, J., & Musche, M. (2023). Livestock density affects species richness and community composition of butterflies: A nationwide study. Ecological Indicators, 146, 109866. https://doi.org/10.1016/j.ecolind.2023.109866
Keerthika, A., & Parthiban, K. T. (2022). Multifunctional agroforestry landscapes: Augmenting butterfly biodiversity at foot hills of Nilgiris, India. International Journal of Tropical Insect Science, 42(1), 545-556. https://doi.org/10.1007/s42690-021-00570-z
Kremen, C. R. K., Colwell, T.L., Erwin. D.D., Murphy, R.F., Noss, R. & Saujayan, M. (1993). Terrestrial arthropod assemblages: their use as indicators in conservation planning. Conservation Biology 7(4): 796-808.
Kurylo, J. S., Threlfall, C. G., Parris, K. M., Ossola, A., Williams, N. S. G., & Evans, K. L. (2020). Butterfly richness and abundance along a gradient of imperviousness and the importance of matrix quality. Ecological Applications: A Publication of the Ecological Society of America, 30(7). https://doi.org/10.1002/eap.2144
Lamas, G. (1976). Notas sobre mariposas peruanas (LEPIDOPTERA). III. Sobre una colección efectuada en el departamento de Tumbes. Revista Peruana de Entomología, 19(1), 8-12.
Lamas, G. (1985). Los Papilionoidea (Lepidoptera) en la Zona Reservada de Tambopata, Madre de Dios, Peru. I: Papilionidae, Pieridae y Nymphalidae (en parte). Revista Peruana de Entomología, 27, 59-73.
Ma, X., Wu, H., Jiang, W., Ma, Y., & Ma, Y. (2015). Interference between Redroot Pigweed (Amaranthus retroflexus L.) and Cotton (Gossypium hirsutum L.): growth analysis. PLoS ONE, 10(6) e0130475, 1-18.
Medina J. (2009). Guía didáctica. Las Mariposas. Granada, Parque de las Ciencias.
Neal, W., Araya, Y., & Wheeler, P. M. (2024). Influence of canopy structural complexity on urban woodland butterfly species richness. Journal of Insect Conservation. https://doi.org/10.1007/s10841-024-00594-z
Nicholls, C. I., & Altieri, M. A. (2013). Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agronomy for Sustainable Development, 33(2), 257–274.
Ospina, I., & Reinoso, G. (2009). Mariposas diurnas (Lepidoptera: Papilionoidea y Hesperioidea) del jardín botánico Alejandro von Humboldt de la Universidad del Tolima (Ibagué – Colombia). Revista Tumbaga, 1(4), 135-148.
Peña, L. E., & Ugarte, A. J. (1996). Las mariposas de Chile (The butterflies of Chile). Santiago de Chile, Editorial Universitaria.
Robbins, R. K., & Nicolay, S. S. (2002). An overview of Strymon Hübner (Lycaenidae: Theclinae: Eumaeini). Journal of the Lepidopterists’ Society, 55(3), 85–100.
Salinas-Gutiérrez, J. L., Méndez, C., Barrios, M., Pozo, C., & Llorente-Bousquets, J. (2009). Hacia una Síntesis de los Papilionoidea (Insecta: Lepidoptera) de Guatemala con una reseña histórica. Caldasia, 31(2), 407-440.
Sánchez-Tlacuahuac, N., Pimentel-Equihua, J. L., Espinosa-Hernández, V., & Vibrans, H. (2023). What do monarchs feed on in winter? nectar sources at hibernation sites. Journal of Insect Conservation, 27(1), 181-191. https://doi.org/10.1007/s10841-022-00433-z
Servicio Nacional de Áreas Protegidas por el Estado (SERNANP). (2019). Sistema de áreas naturales protegidas del Perú. Lima, Ministerio del Ambiente.
Settele, J., Kudrna, O., Harpk,e A., Kühn, I., van Swaay, C., Verovnik, R., Warren, M., Wiemers, M., Hanspach, J., Hickler, T., Kühn, E., van Halder, I., Veling, K., Vliegenthart, A., Wynhof, I., & Schweiger, O. (2008) Climate risk atlas of European Butterfies. Biorisk 1(Special Issue), 1–710. https://doi.org/10.3897/biorisk.1
Sevilleja, C. G., Collins, S., Warren, M. S., Wynhoff, I., Van Swaay, C. A. M., Dennis, E. B. et al. (2020). Assessing butterflies in Europe-European butterfly monitoring schemes-network development: technical report. Wageningen (Netherlands): Butterfly Conservation Europe and ABLE/eBMS.
Shuey, J.A., Meerman, J., Labus, P. et al. (2024). Butterfly community structure in Belize: species richness patterns reveal the role of climate gradients in shaping butterfly community structure and strategies for long-term conservation. J Insect Conserv, 28, 233–249. https://doi.org/10.1007/s10841-023-00535-2
Tassin de Montaigu, C., & Goulson, D. (2024). Factors influencing butterfly and bumblebee richness and abundance in gardens. The Science of the Total Environment, 908, 167995. https://doi.org/10.1016/j.scitotenv.2023.167995
Thomas, J. A. (2005). Monitoring change in the abundance and distribution of insects using butterflies and other indicator groups. Philos Trans R Soc Lond B, 360, 339–357.
Thomas, J. A., Telfer, M.G., Roy, D.B., Preston, C.D., Greenwood, J., Asher, J., Fox, R., Clarke, R.T., & Lawton, J. H. (2004). Comparative losses of British butterflies, birds, and plants and the global extinction crisis. Science, 303(5665), 1879–1881.
Tobar, L. D., Rangel, C. H. J. O., & Andrade-C, M. G. (2001). Las cargas polínicas en las mariposas (Lepidoptera: Rhopalocera) de la parte alta de la cuenca del río El Roble-Quindío-Colombia. Caldasia, 23(2), 549-557.
Van Swaay, C., Regan, E., Ling, M., Bozhinovska, E., Fernandez, M., Marini-Filho, O. J., Huertas, B., Phon, C. K., Korosi, A., Meerman, J., Peer, G., Uehara-Prado, M., Safian, S., Sam, L., Shuey, J.,Taron, D., Terblanche, R., & Underhill, L. (2015). Guidelines for standardized global butterfly monitoring. Group on Earth Observations Biodiversity Observation Network, Lei-pzig, Alemania. GEO BON Technical Series 1, 32 p.
Vargas, H.A., & Duarte, M. (2016). First host plant record for Strymon davara (Hewitson) (Lepidoptera, Lycaenidae) in the highly human –modified coastal valleys of the Atacama Desert. Revista Brasileira de Entomología, 60(4), 352-355.
Vásquez, J., Zárate, R., Huiñapi, P., Pinedo, P., Ramírez, J., Lamas, G. & Vela, P. (2017). Plantas alimenticias de 19 especies de mariposas diurnas (Lepidoptera) en Loreto, Perú. Revista Peruana de Biología, 24(1), 35-42. https://dx.doi.org/10.15381/rpb.v24i1.13109.
Wang, W. L., Suman, D. O., Zhang, H. H., Xu, Z. B., Ma, F. Z., Hu, S. J. Butterfly conservation in China: From science to action. Insects, 11, 661.
Warren, M. S., Maes, D., van Swaay, C. A. M., Goffart, P., Dyck, V., Bourn, N. A. D. et al. (2021). The decline of butterflies in Europe: problems, significance, and possible solutions. Proceedings of the National Academy of Sciences, 118(2), e2002551117.
Willmott, K. R., & Freitas, A. V. L. (2006). Higher level phylogeny of the Ithomiinae (Lepidoptera: Nymphalidae): classification, patterns of larval hostplant colonization and diversification. Cladistics, 22(4), 297-368.
Yu, X.-T., Yang, F.-L., Da, W., Li, Y.-C., Xi, H.-M., Cotton, A.M., Zhang, H.-H., Duan, K., Xu, Z.-B., Gong, Z.-X., Wang, W.-L. & Hu, S.-J. (2023). (2023). Species richness of Papilionidae butterflies (Lepidoptera: Papilionoidea) in the Hengduan Mountains and its future shifts under climate change. Insects, 14(3), 259. https://doi.org/10.3390/insects14030259