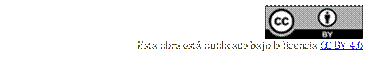
INTRODUCCIÓN
Ocurrencia del hongo Fusarium solani como patógeno causante de la muerte regresiva en el cultivo del aguacate
1 Centro de Investigación Olerícola Frutícola, Universidad Nacional Hermilio Valdizán. Av. Universitaria 601-607, Pillco Marca, Huánuco, Perú.
2 Servicio Nacional de Sanidad Agraria, Huánuco, Perú.
3 Facultad de Ciencias Agrarias, Universidad Nacional Hermilio Valdizán. Av. Universitaria 601-607, Pillco Marca, Huánuco, Perú.
4 Facultad de Ciencias Agrarias, Universidad Nacional Daniel Alcides Carrión, Cerro de Pasco, Perú.
*Autor corresponsal: avalverde@unheval.edu.pe (A. Valverde-Rodríguez).
ORCID de los autores:
N. H. Chavez-Huaman: https://orcid.org/0009-0000-9316-3594 P. Vega y Montecillo: https://orcid.org/0009-0004-5196-339X
L. M. Álvarez Benaute: https://orcid.org/0000-0001-6961-9870 M. J. Castillo Nole: https://orcid.org/0000-0002-4814-8405
M. E. Campos Albornoz: https://orcid.org/0000-0003-0356-9799 A. Valverde-Rodríguez: https://orcid.org/0000-0003-1522-4827
RESUMEN
Palabras clave: aguacate; Fusarium solani; fitopatógenos; cultivo; enfermedades; muerte regresiva.
ABSTRACT
The loss of productive plants due to the dieback of the avocado (Persea Americana Mill) in Peru is of growing concern in the agricultural sector. This devastating phenomenon manifests itself with necrosis in leaves, branches, stems and death of trees. The Fruit and Vegetable Research Center houses 22 varieties of avocado, with the Hass and Fuerte varieties being the most susceptible to the attack of dieback, so the objective of this work Identify the causal agent of dieback in the Hass variety. 20 plants with symptoms of necrosis in leaves, branches and stems were selected, along with other signs such as death of branches, necrosis of the fruit peduncle and development of small fruits from which roots and soil were extracted as a sample. Necrotic roots of plants were collected and processed in the laboratory, which were disinfected, macerated, and sown on PDA agar, isolations, and the respective morphological characterization of the culture on PDA were carried out. The findings made it possible to identify the fungus Fusarium solani as the pathogen responsible for dieback. The confirmation of the phytopathogenic agent was carried out through the report of the National Agrarian Health Service. The pathogenicity of the fungus was demonstrated by inoculating the avocado plants in the nursery and evaluating them six months later, showing symptoms of root rot, scarce and brittle secondary roots. In conclusion, the study demonstrates that Fusarium solani is the pathogen responsible for dieback in the crop.
Keywords: avocado; Fusarium solani; phytopathogens; cultivation; diseases; dieback.
Recibido: 24-06-2024.
Aceptado: 10-11-2024.
INTRODUCCIÓN
El aguacate o palto (Persea americana Miller) de origen Centroamérica y México, ha sido cultivado y consumido desde tiempos antiguos. Su adaptabilidad a diferentes climas y su alto valor nutricional lo han convertido en un cultivo popular en todo el mundo (Valencia et al., 2022). A lo largo de los tiempos, el cultivo del palto ha sido introducido en numerosos países favorecido por su demanda creciente en la industria alimentaria y su reconocido valor gastronómico (Rondón & Vidal, 2020; Bedoya & Julca, 2020). Entre estos países se encuentra Perú, donde ha encontrado condiciones favorables para su cultivo y expansión en los diferentes pisos altitudinales y condiciones climáticas del país, convirtiéndolo a Perú en uno de los principales productores y exportadores de esta fruta y es así como en el 2021 el país cosechó 779 mil toneladas de palta y exportó 526 mil toneladas (Flores-Izquierdo & Espinoza-Villanueva, 2023). La región de la Libertad lidera la producción nacional de aguacate debido a su clima, suelos favorables y proximidad a los puertos costeros, se suma a ello la región Ayacucho, Huancavelica y otros, como es el caso de la región Huánuco, dado a sus condiciones climáticas favorables, en los valles interandinos. Sin embargo, varias enfermedades de rápida propagación afectan la producción de aguacate, entre ellos está la pudrición de la raíz causado por Phytophthora cinnamomi o la recientemente descubierta muerte regresiva provocado por Fusarium euwallaceae, F. kuroshium, F. obliquiseptatum entre otros hongos (Na et al., 2018), y transmitida por dos invasores. Barrenadores (Euwallacea sp. nr. fornicatus y E. kuroshio, también conocidos como barrenadores polífagos (Eskalen et al., 2012; Na et al., 2018; Guevara-Avendaño et al., 2019; Callaghan et al., 2024)., además se ha descrito que el Fusarium solani, es uno de los principales grupos de simbiontes fúngicos de los escarabajos del grupo de Euwallacea (Coleoptera: Curculionidae: Scolytinae) barrenadores polífagos que junto con los mutualistas fúngicos F. ambrosium y F. rekanum, son una plaga invasora en los árboles de aguacate (P. americana) en Florida de Estados Unidos (Paap et al., 2018; Van Rooyen et al., 2021; Callaghan et al., 2024).
En el Centro de Investigación de Horticultura El-Kanater en Egipto, se llevó a cabo una investigación exhaustiva centrada en cepas de Fusarium solani, Fusarium moniliforme y Fusarium semitectum previamente aisladas de raíces de plántulas de vivero, las cepas al ser sometido a pruebas de patogenicidad reprodujeron los síntomas de marchitez o "damping off" en plantas de Persea americana Mill con diferentes niveles de agresividad. Comprobándose que F. solani es el hongo más patógeno, seguido por F. moniliforme (Radwan & Hassan, 2018).
Tanto F. solani como Armillaria spp son los agentes causales de una enfermedad de progresión lenta que resulta letal, provocando el secado y la muerte de los árboles de palto o aguacate, atacan y descomponen el cambium y la madera de las raíces principales (Cantú en 2022). El Fusarium solani y Fusarium oxysporum, fueron identificados a través del análisis morfológico y molecular como causante de la podredumbre radicular y declive en los árboles de aguacate en Turquía (Çalış et al., 2024), también se ha descrito que el patógeno podría verse favorecido por el estrés hídrico en los árboles que desencadena trastornos fisiológicos a los árboles; esto, a su vez, facilita la manifestación de enfermedades de la madera al permitir que los patógenos latentes se establezcan y proliferen tanto en el tejido interno como a través de nuevas colonizaciones (Bejarano-Bolívar et al., 2021).
Las plantas afectadas por cancros en las ramas tienen una alta probabilidad de desarrollar estos mismos síntomas en el tronco y ramitas, generando una corteza endurecida o consistente (Na et al., 2018; Guevara-Avendaño et al., 2019). La progresión de la enfermedad conduce a la muerte regresiva de las ramitas más pequeñas, las cuales retienen hojas y frutos marchitos que pueden persistir durante meses en la planta (Bejarano-Bolívar et al., 2021).
Los hongos asociados al microbioma endofítico de los árboles de aguacate son considerados también, patógenos oportunistas, que, en presencia de daños mecánicos en las raíces, estos hongos pueden invadir el árbol. La subfamilia Scolytinae, conocida por su importancia en el ámbito forestal, frutícola y ornamental, incluyendo P. americana, alberga especies de escarabajos xilomicetófagos y filófagos (Pérez-Silva et al., 2015). Los escarabajos xilomice-tófagos tienen la capacidad de transportar esporas de hongos en micangios, estructuras especializadas, e inocular el hongo en el sistema vascular de los árboles vivos. Esta interacción causa la obstrucción del flujo de agua y nutrientes, lo que puede llevar a la muerte parcial o total de la planta en un corto período de tiempo (Harrington et al., 2008). Además, esta simbiosis favorece la dispersión del hongo a nuevos hospederos (García-Avila et al., 2016).
En Perú, Ancistrosoma klugii ha sido registrado como habitante de los suelos cultivados con el aguacate, las larvas se alimentan de las raíces de los árboles en campo y los adultos consumen los follajes (Lupaca, 2015) y al mismo tiempo en las parcelas del Centro de Investigación Olerícola Frutícola de la Universidad Nacional Hermilio Valdizan, se ha registrado problemas de muerte regresiva en las plantas del aguacate, donde hasta la fecha no se cuenta con la información sobre el complejo de patógenos que causan la muerte regresiva. Así, el presente estudio se centra en identificar el o los agentes causales de la enfermedad. Por lo que se aislaron, identificaron y se realizaron pruebas de patogenicidad del agente causal.
METODOLOGÍA
Los estudios se llevaron a cabo en el Centro de Investigación Olerícola Frutícola de la Universidad Nacional Hermilio Valdizan, que custodia 22 variedades del palto con fines experimentales, y el laboratorio de Microbiología de la Facultad de Ciencias Agrarias de la misma universidad, situado geográficamente en las coordenadas 09° 57' 03" O y 77° 14' 79" S, a una altitud de 1947 msnm y el laboratorio de Identificación de Patógenos del Servicio Nacional de Sanidad Agraria (SENASA).
Se ocuparon 20 muestras de raíces entre las variedades Hass y Fuerte para el aislamiento del inóculo en PDA y la respectiva caracterización morfológica e identificación del patógeno en el Laboratorio. Similar cantidad de muestras, tanto de raíces y suelo fueron enviados al laboratorio del Servicio de Sanidad Agraria, para el análisis, identificación y confirmación del patógeno causante de la muerte regresiva.
Fase de campo- muestreo, se examinaron veinte arboles con muerte regresiva (Figura 1), entre las dos variedades que bajo las condiciones estudiadas resultaron demostrar significativamente los síntomas de la enfermedad como amarilleamiento, defoliación, necrosis, declive del follaje y muerte, de estas plantas se extrajeron las raíces, para el aislamiento e identificación del patógeno.
Las raíces colectadas tuvieron clara evidencia de síntomas como la podredumbre seca, de un color oscuro y quebradizos, las muestras fueron sumergidos en una solución de hipoclorito sódico al 2% durante tres minutos para desinfectarlos. A continuación, se lavaron dos veces con agua destilada estéril y se sellaron en bolsas de polietileno.
Fase de laboratorio: Una vez que la estructura fúngica se hizo evidente, las raíces se aislaron en cajas Petri de 90 mm de diámetro utilizando 20 mL de medio de cultivo agar papa dextrosa (PDA). Las cajas se instalaron en una cámara climática a 25 °C, 70% HR, con 12 horas de luz y 12 horas de oscuridad durante quince días. El proceso de elaboración de PDA acidificado fue sugerido por Walker & White (2017).
Pruebas de patogenicidad. Los aislados del patógeno fueron cultivados en PDA a 25 °C 70% R, con 12 horas de luz y 12 horas de oscuridad, durante 21 días. Los conidios se recogieron en agua estéril y se diluyeron en 6 mL. La suspensión de macro y microconidios, estos fueron utilizados para inocular a las plantas del palto libres de enfermedades, de cuatro meses de edad y cultivadas en sustrato estéril, provenientes del vivero frutícola del centro de investigación. Para la inoculación se provocaron heridas a la altura del cuello de la planta, con la ayuda de una navaja estéril y se inocularon las suspensiones de conidios a través de las heridas. Se ocuparon cinco plantas para la prueba de patogenicidad del inóculo y tres plantas sanas como testigo, a los 180 días después de la inoculación del patógeno en las plantas fueron evaluadas la patogenicidad de los aislados.

Figura 1. plantas con síntomas de muerte regresiva.
Caracterización e identificación: Para la caracterización e identificación de los géneros en el laboratorio se utilizaron las claves morfológicas correspondientes (Erwin & Ribeíro, 1996) y el microscopio Microscopio Compuesto BiolóGico, Lente objetivo: 4X 6X 10X 60X, Binocular Hd, Iluminación Led Dual.
RESULTADOS Y DISCUSIÓN
Descripción de las características morfológicas del hongo
A los 8, 15 y 21 días de evaluación, el color de la colonia y la forma de las macroconidias fueron las diferencias más notables observadas. Tras aislar el hongo y cultivarlo durante ocho días, la tasa de crecimiento radial medida en ese momento fue de 1,6 cm, lo que indica una tasa media de crecimiento de 7,33 mm/día. Las colonias aparecieron inicialmente como micelio blanco y velloso, que al transcurso de los días se tornaron a un color púrpura tenue. Micelio sin formación de esporodoquios hasta este momento, placas observadas al reverso de colonia con coloración naranja tenue.
En 5 días de cultivo fue posible visibilizar la pigmentación purpura en el punto de siembra del hongo, con formación de esporodoquios, masas de conidióforos cortos y estrechamente ramificados que emergen rectos de una maraña de hifas, rectos y robustos en su gran mayoría y un mínimo porcentaje de micelio en colonias ligeramente curvos. Observados al reverso de la placa, clamidosporas en ubicación intercalados o terminales de formas esféricas y de paredes rugosas de tonalidades beige – naranjo (Figura 2 A-B) observadas al reverso de colonia con coloración naranja tenue.
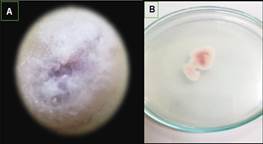
Figura 2. Características morfológicas de Fusarium solani. (A) colonia de 6 días en PDA, con micelio aéreo blanco algodonoso (B) Fusarium solani
A los 15 días del cultivo fue posible visibilizar la pigmentación purpura en el punto de siembra del hongo, con formación de esporodoquios, masas de conidióforos cortos y estrechamente ramificados que emergen de las hifas, rectos y robustos en su gran mayoría y un mínimo porcentaje de micelio en colonias ligeramente curvos. Observados al reverso de la placa, clamidosporas en ubicación intercalados o terminales de formas esféricas y de paredes rugosas de tonalidades beige – naranjo (Figura 3).
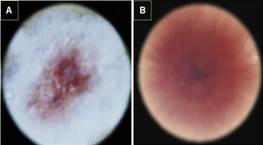
Figura 3. Características morfológicas de los aislamientos de Fusarium solani en PDA. A) Color de la colonia en PDA en 15 días B) Observados al reverso de la placa de tonalidades beige – naranjo.
A los 21 días de evaluación fue significativo el crecimiento radial con 2,6 cm, crecimiento micelial algodonoso al principio blanca a verde para luego convertirse a blanco-naranjo con diferenciación de microconidióforos hialinos septados de tipo fusiforme (Figura 4), con 5-6 septos. Microconidias ovales o elipsoidales, reniformes o redondeados solitarios no septados de colores verdes amarillen-tas, crema o hialinas y las macroconidias común-mente 4 septos, de formas curvos dorsalmente, algunos elongados de color crema en las etapas del desarrollo, para tornarse a naranjo al final, estas características confirmaron la presencia del patógeno Fusarium solani en los árboles de aguacate en la zona de estudio.
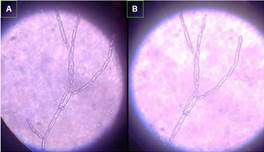
Figura 4. Microconidióforos hialinos septados de tipo fusiforme.
Forma de las colonias
Colonias de forma pionnotes con bordes lizos y capa superficial de apariencia algodonosa. En el otro lado de la placa de Petri, se observó un tono blanco más claro cerca del borde y un color naranja más oscuro hacia el centro., colonias de forma irregular oval, micelio de color naranjo oscuro en PDA a los 21 días de desarrollo. Los macroconidios de forma fusoide desigual (Figura 5).
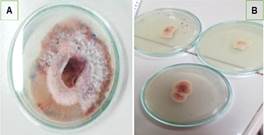
Figura 5. A) Colonia de Fusarium solani de forma pionnotes, capa de apariencia algodonosa. B) Crecimiento y coloración de la colonia vista por el reverso de la caja de Petri.
Identificación de la especie fitopatógena causante de la muerte regresiva en el cultivo de palto en CIFO-UNHEVAL con resultado del laboratorio SENASA.
Clasificación taxonómica
División: Eumycota,
Subdivisión: Deuteramycotina,
Clase: Hiphomycetes,
Orden: Moniliales,
Familia: Tuberculariaceae,
Género: Fusarium,
Especie: Fusarium solani
Identificación del agente causal
|
N° |
Tipo de muestra Persea americana |
Raíz 20 Unds primera muestra |
|
|
Resultados |
|
|
1 |
Positivo a la presencia de |
Fusarium solani |
Fuente: Sistema de Vigilancia Fitosanitaria, SENASA, diciembre 2022.
Patogenicidad del inóculo en las plantas de palto a nivel de vivero
A los 6 meses después de la inoculación en las plantas de palto a nivel de vivero, no fue posible evidenciar los síntomas de marchites en las plantas inoculadas con la suspensión de Fusarium solani, sin embargo, se pudo observar en las hojas basales los síntomas de amarillamiento y quemaduras, tamaño de los tallos de entre 37 a 48 cm. Las raíces de plantas inoculadas con pudrición y de aspecto ennegrecido y al romperse los tallos la evidencia de manchas de color castaño-rojizo en la línea de los vasos conductores. Generalmente raíces escasas y quebradizas, de menor tamaño, de entre 12 a 28 cm, color marrón oscuro; y al corte de la base del cuello y la raíz principal, el tejido leñoso muestra una coloración marrón, típica de la podredumbre radicular, y la raíz principal con decoloración y necrosis por F. solani (Figura 6). Estos síntomas son parecidos a lo reportado por Carranza Rojas et al. (2015) al realizar le aislamiento, identificación y patogenicidad de hongos asociados a la tristeza del aguacatero en México, donde la frecuencia de la ocurrencia del patógeno F. solani fue de hasta 10,8%., junto con otros patógenos que formaban el complejo de hongos, los autores a la vez señalaron que las cepas de Fusarium sp. y P. cinnamomi fueron las de mayor severidad en los daños al tejido en la raíz.
|
|
|
|
|
|
|
|
|
Figura 6. Izquierdo, tallos y hoja de plantas inoculadas con F. solani. Derecha, raíces de plantas inoculadas con F. solani, imagen a los 8 meses de tratamiento.
El grupo testigo, plantas sin inocular evidenciaron menor tamaño de los tallos, de hasta 24 cm y mayor crecimiento de raíces de entre 37 cm a 44 cm. Mejor desarrollo de raíces secundarios, raíces de color marrón claro, no quebradizas. En tanto las plantas inoculadas meses después de la inoculación a nivel de vivero, manifestaron mayor vigorosidad y crecimiento, hojas apicales lozanas y verde brillante, con raíces deformes, escaso desarrollo y quebradizos (Figura 7).
En campo las plantas establecidas manifestaron síntomas de muerte regresiva, cancros en las ramas y tallos apicales, estas sintomatologías tienen cierta similitud con los descritos por Koyyappurath et al., (2015) al describir los síntomas causados por Fusarium spp. en los aguacateros, como la abscisión precoz de las hojas, las hojas pequeñas, el amarilleamiento de las hojas y la interrupción del crecimiento vegetativo de los brotes. Además, existe una conexión entre estos rasgos y la descripción por Pegg et al. (2007) al evaluar las enfermedades foliares, frutales y del suelo en el cultivo del palto.
|
|
|
Figura 7. Izquierda (plantas libres de F. solani), Derecha (Plantas inoculada con F. solani), notoria diferencia en el desarrollo de las raíces y la parte vegetativa.
En PDA, el crecimiento lento para llegar hasta un promedio 2,6 cm, crecimiento micelial algodonoso al principio blanca a verde para luego convertirse a blanco-naranjo con diferenciación de microco-nidióforos hialinos septados de tipo fusiforme concuerdan con los reportados por Chinchilla-Salazar et al. (2020) para la especie F. solani. Las características más distintivas de las especies de Fusarium spp. en las zonas muestreadas relacionadas con la marchitez vascular, según la identificación molecular y morfológica, fueron la forma de los microconidios, el número de septos, el color de los esporodoquios y la forma de los macroconidios. Gregorio et al. (2020) al caracte-rizar el Fusarium spp., como fitopatógeno en el cultivo del palto en México después de una incubación durante 7 días, informó que los micelios en la etapa temprana de crecimiento son de color blanco para luego convertirse a marrón pálido en la etapa tardía de crecimiento por su parte Jeon et al. (2013) describieron que los aislados de F. solani produjeron abundantes microconidios.
El tiempo transcurrido hasta que aparecieron los primeros síntomas aéreos fluctuó entre 3 a 6 meses después de la inoculación del Fusarium, al respecto Berruezo (2018) al caracterizar morfológica, biológica y molecularmente los complejos F. oxysporum y F. solani asociados al cultivo de Nicotiana tabacum L. en el Noroeste Argentino, observó que F. solani incrementó la severidad a mayor concentración de inóculo, la incidencia de los síntomas de marchitamiento aumentaba lineal-mente con el aumento de la densidad de inóculo. Esta característica también es descrita por Radwan & Hassan (2018) quienes realizaron pruebas de patogenicidad en semillas y plántulas del palto en condiciones de invernadero, registrando a F. solani fue el hongo más patógeno que causó el mayor porcentaje de pudrición de raíces pre y post-emergente. Este reporte es similar a los descrito por Gregorio et al. (2020) al caracterizar el Fusarium spp., como fitopatógeno en el cultivo del palto en México., quienes indican que las plantas inoculadas reflejaron un crecimiento reducido de la planta, así como una coloración marrón rojiza en su tejido indicando necrosis, raíces de coloración rojo pardusco. Parra-Cota et al. (2019) reportaron a F. solani como un patógeno secundario en la pudrición de la raíz de los cítricos, dado que a corto plazo no es capaz de causar la muerte de la planta, a pesar de colonizar gran parte de la raíz, sin embargo, reduce el vigor de la planta; esta afirmación validó a lo señala por Olalde-Lira (2020) reportó que F. solani produjo marchitez, defoliación y posterior muerte de la planta al causar la pudrición de la raíz a nivel de vivero; lo que difiere de lo informado por Gaetán (2007) donde F. solani produjo pudrición de la raíz y la corona dos semanas después de la inoculación. Koyyappurath et al. (2015) sus obser-vaciones de la patogenicidad del Fusarium spp incluyeron la pudrición de raíces pequeñas con una tonalidad marrón-negra oscura que, con el tiempo, se extiende a las raíces más gruesas.
CONCLUSIONES
Dado el impacto de Fusarium solani en el desarrollo y productividad de los cultivos, se recomienda realizar estudios futuros que profundicen en estrategias de manejo biológico y control integrado del patógeno, así como en la identificación de variedades de cultivos con resistencia natural, lo cual permitirá implementar prácticas más efectivas y sostenibles para mitigar su propagación y daño.
AGRADECIMIENTOS
El Director del Centro de Investigación Frutícola Olerícola-UNHEVAL, Huánuco, por haber contribuido con todo el apoyo logístico para la toma de muestras en las parcelas del banco de germoplasma del palto.
REFERENCIAS BIBLIOGRÁFICAS
Bedoya Justo, E., & Julca Otiniano, A. (2020). Caracterización de fincas productoras del cultivo de palto en la región Moquegua, Perú. Idesia (Arica), 38(3), 59-67. http://dx.doi.org/10.4067/S0718-34292020000300059
Bejarano-Bolívar, A. A., Lamelas, A., von Wobeser, E. A., Sánchez-Rangel, D., Méndez-Bravo, A., Eskalen, A., & Reverchon, F. (2021). Shifts in the structure of rhizosphere bacterial communities of avocado after Fusarium dieback. Rhizosphere, 18, 100333. https://doi.org/10.1016/j.rhisph.2021.100333
Berruezo, L. A. (2018). Caracterización morfológica, biológica y molecular de los complejos Fusarium oxysporum y Fusarium solani asociados al cultivo de Nicotiana tabacum L. en el Noroeste Argentino.
Çalış, Ö., Çelik, S., Fidan, H., Tek, M. I., Shah, M., Tozlu, I., & Wani, S. H. (2024). Emerging pathogens and disease dynamics threatening avocado production in southern Türkiye. Journal of Plant Diseases and Protection, 1-11. https://doi.org/10.1007/s41348-024-00954-6
Callaghan, S., Carnegie, A. J., Gillespie, P., Mulholland, S., Nagel, M., Sargeant, D., ... & Wildman, O. (2024). Response to the detection of Fusarium dieback associated with ambrosia beetles on Acer negundo in New South Wales. Australasian plant pathology, 53(4), 345-352. https://doi.org/10.1007/s13313-024-00984-6
Cantú Treviño, K. G. (2022). Etiología de la muerte regresiva de árboles de aguacate en Sabinas Hidalgo, Nuevo León (Doctoral dissertation, Universidad
Carranza Rojas, Y., Morales García, J. L, Pedraza Santos, M. E., Chávez Bárcenas, A. T., & Morales Montelongo, K. L. (2015). Aislamiento, identificación y patogenicidad de hongos asociados a la tristeza del aguacatero en Michoacán, México. Plagas y Enfermedades, 189–193. https://doi.org/10.22533/at.ed.9732172210124
Chaupín Mejía, M. O. (2018). Incidencia, etiología y control in vitro de la muerte regresiva en el palto (Persea americana Mill.) en Luricocha, Huanta, 2017.
Chinchilla-Salazar, G., Blanco-Meneses, M., & Castro-Zúñiga, Ó. (2020). Identificación molecular y morfológica de las especies de fusarium spp., asociadas al cultivo de pimienta negra (Piper nigrum) en Sarapiquí y Guatuso en Costa Rica. Agronomía Costarricense, 44(2), 1-30.
Erwin, D., & Ribeiro, O. (1996). Phytophthora cinnamomi var. cinnamomi. The American Phytopathological Society, pp. 269-272.
Eskalen, A., González, A., Wang, D. H., Twizeyimana, M., Mayorquin, J. S., & Lynch, SC (2012). Primer reporte de un Fusarium sp. y su vector barrenador del té (Euwallacea fornicatus), que causa la muerte regresiva por Fusarium en el aguacate en California. Enfermedad de las plantas, 96(7), 1070-1070. https://doi.org/10.1094/PDIS-03-12-0276-PDN
Flores-Izquierdo, M. A., & Espinoza-Villanueva, L. E. (2023). Situación actual y perspectivas de la producción de palta (Persea americana) peruana en el contexto del comercio internacional. Ingeniería Industrial, 45, 1-17. https://doi.org/10.26439/ing.ind2023.n45.6316
Gaetán, S. A., Madia, M. S., & Perez, A. (2007). Recent outbreak of Fusarium crown and root rot caused by Fusarium solani on marjoram in Argentina. Australasian Plant Disease Notes, 2(1), 15-16. https://doi.org/10.1071/DN07006
García-Avila, C. D. J., Trujillo-Arriaga, F. J., López-Buenfil, J. A., González-Gómez, R., Carrillo, D., Cruz, L. F., ... & Acevedo-Reyes, N. (2016). First report of Euwallacea nr. fornicatus (Coleoptera: Curculionidae) in Mexico. Florida Entomologist, 99(3), 555-556. https://doi.org/10.1653/024.099.0335
García-Guevara, J. F., García-Ávila, C. D. J., Acevedo-Reyes, N., & Vergara-Pineda, S. (2018). Escarabajos (Curculionidae: Scolytinae) Asociados a trampas en huertos de Persea americana Miller, 1768 en cuatro municipios de Michoacán. Entomologia mexicana, 5, 408-414.
Gregorio Olalde-Lira, G., Raya Montano, Y. A., Apaez Barrios, P., Vargas-Sandoval, M., Pedraza Santos, M. E., Raymundo, T., ... & Nieves Lara-Chavez, M. B. (2020). Characterization of Fusarium spp., a Phytopathogen of avocado (Persea americana Miller var. drymifolia (Schltdl. and Cham.)) in Michoacan, Mexico. Revista de la facultad de ciencias agrarias, 52(2), 301-316.
Guevara-Avendaño, E., Bejarano-Bolívar, A. A., Kiel-Martínez, A. L., Ramírez-Vázquez, M., Méndez-Bravo, A., von Wobeser, E. A., ... & Reverchon, F. (2019). Avocado rhizobacteria emit volatile organic compounds with antifungal activity against Fusarium solani, Fusarium sp. associated with Kuroshio shot hole borer, and Colletotrichum gloeosporioides. Microbiological research, 219, 74-83. https://doi.org/10.1016/j.micres.2018.11.009
Harrington, T. C., Fraedrich, S. W., & Aghayeva, D. N. (2008). Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the Lauraceae. Mycotaxon, 104(2), 399-404.
Jeon, C. S., Kim, G. H., Son, K. I., Hur, J. S., Jeon, K. S., Yoon, J. H., & Koh, Y. J. (2013). Root rot of balloon flower (Platycodon grandiflorum) caused by Fusarium solani and Fusarium oxysporum. The plant pathology journal, 29(4), 440.
Koyyappurath, S., Atuahiva, T., Guen, R. Le, Batina, H., Squin, S. L., Gautheron, N., Edel Hermann V., Peribe, J., Jahiel, M., Steinberg, C., Liew, E. C., Alabouvette, C., Beese, P., Dron, M., Sache, I., Laval, V., Grisoni, M. (2015). Fusarium oxysporum f. sp. radicis-vanillae is the causal agent of root and stem rot of vanilla. Plant Pathology. 1-14. https://doi.org/10.1111/ppa.12445
Llauger, E. R. (2016). La fruticultura y el desarrollo sostenible. FAO. Sitio web: http://www.fao.org/cuba/noticias/detail-events/en/c/412715/
Lupaca, D. A. (2015). Control del ragao en el valle de Huánuco. Investigación Valdizana, 9(1), 4-9.
Morelos, E. M., Lino, M. D., Ortega, E., & Castellanos, P. L. (2021). Evaluación de la capacidad antagónica in vitro de cepas de Trichoderma spp frente a Phytophthora cinnamomi, fitopatógeno de Persea americana (Palta). Ciencia e Investigación, 23(1), 65-70.
Na, F., Carrillo, J. D., Mayorquin, J. S., Ndinga-Muniania, C., Stajich, J. E., Stouthamer, R., ... & Eskalen, A. (2018). Two novel fungal symbionts Fusarium kuroshium sp. nov. and Graphium kuroshium sp. nov. of Kuroshio shot hole borer (Euwallacea sp. nr. fornicatus) cause Fusarium dieback on woody host species in California. Plant Disease, 102(6), 1154-1164. https://doi.org/10.1094/PDIS-07-17-1042-RE
Olalde-Lira, G. G., Raya Montaño, Y. A., Apáez Barrios, P., Vargas-Sandoval, M., Pedraza Santos, M. E., Raymundo, T., ... & Blanca, M. (2020). Caracterización de Fusarium spp., fitopatógeno de aguacate (Persea americana Miller var. drymifolia (Schltdl. y Cham.)) en Michoacán, México. Revista de la Facultad de Ciencias Agrarias. Rev. FCA UNCUYO, 52(2), 301-316.
Paap, T., De Beer, Z. W., Migliorini, D., Nel, W. J., & Wingfield, M. J. (2018). The polyphagous shot hole borer (PSHB) and its fungal symbiont Fusarium euwallaceae: a new invasion in South Africa. Australasian Plant Pathology, 47, 231-237. https://doi.org/10.1007/s13313-018-0545-0
Parra-Cota, F. I., García Pereyra, J., Aviña Martínez, G. N., & Santos-Villalobos, S. D. L. (2019). Primer reporte de marchitamiento por Fusarium en Citrus sinensis var. Valencia en el Valle del Yaqui, México. Revista mexicana de fitopatología, 37(1), 193-201. https://doi.org/10.18781/r.mex.fit.1810-3
Pegg, K. G; Coates, L. L; Corsten, L.; & Harding, R. M. (2007). Enfermedades foliares, frutales y del suelo. En: Whiley, AW; Schaffer, B. y Wolstenholme, BN 2007. El palto: botánica, producción y usos. Traducción. Ediciones Universidad de Valparaíso. Litogarina. Valparaíso. pag. 275-3.
Pérez Álvarez, S., Ávila Quezada, G., & Coto Arbelo, O. (2015). El aguacatero (Molino de Persea americana). Cultivos Tropicales, 36(2), 111-123.
Pérez-Silva, M., Equihua Martínez, A., Estrada Venegas, E. G., Muñoz Viveros, A. L., Valdez Carrasco, J. M., Sánchez Escudero, J., & Atkinson, T. H. (2015). Synopsis of Mexican species of the genus Xyleborus Eichhoff, 1864 (Coleoptera: Curculionidae: Scolytinae). Acta zoológica mexicana, 31(2), 239-250.
Radwan, M. A., & Hassan, M. S. S. (2018). Control of Avocado (Persea americana Miller) Damping-off and Root Rot Diseases in Egypt. Journal of Plant Protection and Pathology, 9(9), 577-585.
Rondón, G. L. F., & Vidal, L. A. G. (2020). Análisis de competitividad para cultivo de palta peruana. Periodo 2000 a 2017. Anales Científicos, 81(1), 58-70. https://doi.org/10.21704/ac.v81i1.1554
Valencia, A. L., Saavedra-Torrico, J., Rosales, I. M., Mártiz, J., Retamales, A., Link, A., & Gil, P. M. (2022). Unveiling the predisposing factors for the development of branch canker and dieback in avocado: a case of study in Chilean orchards. Horticulturae, 8(12). https://doi.org/10.3390/horticulturae8121121
Van Rooyen, E., Paap, T., De Beer, W., Townsend, G., Fell, S., Nel, W. J., ... & Roets, F. (2021). The polyphagous shot hole borer beetle: Current status of a perfect invader in South Africa. South African Journal of Science, 117(11-12), 1-10. https://doi.org/10.17159/sajs.2021/9736
Walker, G. M., & White, N. A. (2017). Introduction to fungal physiology. Fungi: biology and applications, 1-35. https://doi.org/10.1002/9781119374312.ch1.







