
INTRODUCCIÓN
Identificación y caracterización morfológica de endoparásitos en corvina (Plagioscion squamosissimus)
Identification and morphological characterization of endoparasites in Silver Croaker (Plagioscion squamosissimus)
Jenny Virhuez R.1; Victor Puicón N.2*; German Augusto Murrieta Morey3, 4
1 Universidad Peruana Cayetano Heredia, Lima, Perú.
2 Grupo de investigación de Parasitología Veterinaria y Zoonosis Parasitaria, Facultad de Medicina Veterinaria, Escuela Profesional de Medicina Veterinaria, Universidad Nacional de San Martín. San Martín, Perú.
3 Instituto de Investigaciones de la Amazonía Peruana (IIAP). Laboratorio de Parasitología y Sanidad Acuícola, Loreto, Perú.
4 Universidade Estadual do Maranhão (UEMA). Programa de Pós graduação em Ciência Animal (PPGCA), Maranhão, Brazil.
* Autor corresponsal: vhpuicon@unsm.edu.pe (V. Puicón).
ORCID de los autores:
J. Virhuez R.: https://orcid.org/0009-0005-8758-3919 V. Puicón N.: http://orcid.org/0000-0003-2532-2551
G. A. Murrieta Morey: http://orcid.org/0000-0001-6244-2654
RESUMEN
El objetivo del presente estudio fue identificar los taxones de la fauna endoparasitaria, así como su frecuencia, intensidad, abundancia y órganos afectados en Plagioscion squamosissimus (corvina) procedentes de cuatro mercados de la ciudad de Iquitos – Perú. Un total de 60 ejemplares de P. squamosissimus fueron examinados a través de la observación, búsqueda e identificación directa. Se estudiaron los ojos, piel, músculo y vísceras en búsqueda de endoparásitos. Se identificaron larvas de Nematoda correspondientes a: Hysterothylacium sp. y Spiroxys sp. en ojos, Contracaecum sp. y Procamallanus (Spirocamallanus) inopinatus (adulto) en intestino, Anisakis larva tipo 1 y Pseudoterranova sp. en cavidad celómica e intestino, y una especie de acantocéfalo Rhadinorhynchus plagioscionis en ciegos pilóricos. Así mismo se realizó el cálculo de los índices parasitarios (frecuencia, intensidad, intensidad media y abundancia media) y la correlación del tamaño y peso de los peces con la abundancia parasitaria. Se concluye que la fauna endoparasitaria presente en P. squamosissimus está compuesta en su mayoría por especies de nematodos, existiendo una alta diversidad de especies en ejemplares provenientes del medio natural.
Palabras clave: Acantocéfalos; Amazonía Peruana; Anisakidae; endoparásitos; nemátodos.
ABSTRACT
The objective of the present study was to identify the taxa of the endoparasitic fauna, as well as their frequency, intensity, abundance and affected organs in Plagioscion squamosissimus (Silver croaker) from four markets in the city of Iquitos - Peru. Sixty specimens of P. squamosissimus were examined through observation, search, and direct identification. Eyes, skin, muscle, and viscera were studied for endoparasites. Nematoda larvae were identified corresponding to: Hysterothylacium sp. and Spiroxys sp. in eyes, Contracaecum sp. and Procamallanus (Spirocamallanus) inopinatus (adult) in intestine, Anisakis larva type 1 and Pseudoterranova sp. in coelomic cavity and intestine, and a species of acanthocephalan Rhadinorhynchus plagioscionis in pyloric caecae. Parasitic indices (frequency, intensity, mean intensity and mean abundance) and the correlation of fish size and weight with parasitic abundance were also calculated. It is concluded that the endoparasitic fauna present in P. squamosissimus is mostly composed of nematode species, with a high diversity of species in specimens from the natural environment.
Keywords: Acanthocephalans; Anisakidae; endoparasites; nematodes; Peruvian Amazon.
Recibido: 10-09-2024.
Aceptado: 10-12-2024.
INTRODUCCIÓN
La mayor diversidad y abundancia de peces del mundo se encuentra en la Amazonía (Galvis et al., 2006; García et al., 2018). Se estima que a nivel amazónico existen de 2,500 a 3,000 especies de peces, encontrándose en Perú unas 1,200 especies aproximadamente, varios de ellos son cruciales para la supervivencia económica y nutricional de la población amazónica (García et al., 2018; Murrieta, 2019). Se conoce que 79 especies de peces son comercializadas en los mercados de Loreto, Ucayali y Madre de Dios (García et al., 2018). Estas repre-sentan alrededor de 80,000 toneladas anuales de desembarque con un valor económico superior a $80 millones (García et al., 2018; Tello et al., 2007).
La pesquería amazónica, por lo tanto, posee una especial importancia en la economía regional, que contribuye a su vez en el PBI nacional, a la oferta de empleo y a la generación de ingresos en la población. Asimismo, presenta importancia social, pues el pescado es la fuente principal de proteínas en la alimentación de la población amazónica (PRODUCE, 2019). Se han estimado que los niveles de consumo llegan a 101 kg per cápita al año en las riberas de los ríos mientras que en las ciudades llegan a 36 kg (García et al., 2018; Tello et al., 2007; PRODUCE, 2019).
Dentro de las especies de peces procedentes de extracción natural; el boquichico (Prochilodus nigricans), doncella (Pseudoplatystoma punctifer), palometa (Mylossoma albiscopum), sábalo (Brycon amazonicus) y corvina (Plagioscion squamosissi-mus) son las de mayor ingreso y consumo en la región de Loreto (Ministerio de la Producción, 2019; Morey, 2019). Plagioscion squamosissimus destaca entre estas especies como un pez importante para la seguridad alimentaria, con una alta disponibilidad de suministro y aceptación por parte de la población debido al agradable sabor de su carne. (García et al., 2018; García-Vásquez et al., 2012). Se estima cifras de 229 t/año en la producción pesquera continental de la amazonia peruana. Loreto reporta los mayores volúmenes de desembarque con promedios entre los 100 a 145 t/año, Ucayali con un promedio de 41 t/año y Madre de Dios, con capturas reducidas, no superando las 6 t/año (García et al., 2018; Tello et al., 2007, PRODUCE, 2019).
La extracción total de peces amazónicos, incluyendo a la corvina, es destinada al consumo humano directo -fresco, congelado o curado- (Morey, 2019; PRODUCE, 2019). Es allí donde radica la importancia del cumplimiento de los requerimientos sanitarios nacionales e internacionales para asegurar la inocuidad de estos alimentos, garantizando así una alimentación libre de patógenos como los parásitos y alta en proteínas (Eiras et al., 2010; Serrano-Martínez et al., 2017). A pesar de la elevada frecuencia de diversos parásitos en los peces amazónicos, pocos estudios se han centrado en su diversidad, sobre todo en los nematodos que afectan a los peces de agua dulce. (Eiras et al., 2010). El parasitismo en peces provoca pérdidas socioeconómicas, no solo por el deterioro o daño que puede causar al producto, sino también por la posibilidad de transmisión de parásitos de importancia en salud pública, cuyos productos deben ser descartados según lo establece la nor-mativa sanitaria vigente (Serrano-Martínez et al., 2017).
La población iquiteña ha desarrollado una prefe-rencia cada vez mayor hacia el consumo de peces crudos o poco cocidos, ya sea bajo la preparación del popular plato nacional, el ceviche o platos de origen oriental como sushi (Brack, 1997; Matta, 2014; García et al., 2018). Por lo tanto, es impor-tante generar estudios sobre las especies de endo-parásitos existentes en los peces comercializados en la ciudad de Iquitos y reportes sobre los problemas en salud pública por el consumo de pescado crudo contaminado, las cuales son muy limitadas o inexistentes (Eiras et al., 2010; Morey, 2019). La diversidad parasitaria que afecta a P. squamosissimus está directamente relacionada a los hábitos alimenticios de la especie, ya que consume moluscos acuáticos, insectos, copépodos, crustáceos y peces, la mayoría de ellos hospederos intermedios o paraténicos de varios parásitos, afectando su estado sanitario (Takemoto et al., 2009).
A pesar de la creciente importancia económica que está presentando P. squamosissimus entre las principales especies de extracción y consumo en la Amazonía peruana (Ministerio de la Producción, 2019), hasta la fecha se han realizado pocos estudios sobre la fauna parasitaria de esta región, y la investigación se ha centrado sobre todo en conocer su biología y su descripción taxonómica. (Ricker, 1975).
En el 2012, en tres cuencas de Brasil, se realizó una investigación de tipo cualitativa con ejemplares de P. squamosissimus como población de estudio, fueron colectados y examinados 105 especímenes, los cuales fueron medidos y pesados. En la investigación reportaron la presencia de parásitos zoonóticos como Austrodiplostomum sp. (larvas), quistes de cestodos, Contracaecum sp. (larvas). Todos los parásitos reportados fueron encontrados en estadio larval y que la correlación negativa entre la condición de los hospederos y la abundancia de parásitos en los lugares de muestreo refleja la patogenicidad de las larvas de nematodos pertenecientes a este orden, que se sabe que causan daños graves a sus huéspedes (Lacerda et al., 2012). En el 2020, en dos cuencas del estado brasileño de Pará, se realizó un estudio cualitativo con P. squamosissimus como población de estudio, la colecta se llevó a cabo en dos puntos de la cuenca colectando un total de 75 especímenes. Se reportó la presencia de Austrodiplostomum compactum (zoonótico), Anisakis sp. (zoonótico), Procama-llanus (Spirocamallanus) sp., Pseudoproleptus sp. Rhadinorhynchus plagioscionis (Acanthocephala), Neoechinorhynchus sp. (Acanthocephala), Ptycho-bothriidae gen. sp. (Cestoda). Se concluyó que se encontraron parásitos en el 97,2% de los peces examinados, existiendo dos parásitos con potencial zoonótico en ejemplares provenientes tanto del río Amazonas como en el Río Tapajós. De un total de 73 ejemplares de P. squamosissimus, los hospedadores exhibieron infección por al menos una especie de parásito, con un predominio de hospedadores infectados por tres y cuatro especies de parásitos (Chagas de Souza et al., 2020).
Los acantocéfalos como Rhadinorhynchus sp., Echinorhynchus sp., Acanthocephalus sp., y Neoechinorynchus sp., se han registrado infectando al atún caballa (Euthynnus affinis) provenientes del puerto de Tangunj Balai, Indonesia (Yusni et al., 2022), el estudio de estos acantocéfalos junto con otros grupos taxonómicos son considerados como centinelas para la contaminación por metales pesados, siendo encontrados en concentraciones más altas en monogeneos y nematodos, teniendo utilidad y aplicación en el cuidado medioambiental (Nachev etc al., 2021).
La mayoría de los ecosistemas tienen una gran variedad de organismos parásitos. Pueden encon-trarse en todos los niveles tróficos y en casi todas las cadenas alimentarias (Morais et al, 2019). La mayoría de los seres vivos del planeta actúan como hospedadores de una o varias especies de parásitos. Además, cada vez se reconoce más que el parasitismo desempeña un papel organizativo crucial en los grupos animales (Lacerda et al., 2012; Morais et al., 2019). A pesar del hecho de que pueden controlar las poblaciones de hospedadores y tener un impacto en la estructura de la comunidad, los parásitos suelen ser ignorados en la investigación sobre las cadenas alimentarias y los ecosistemas (Alves & Luque, 2001; Lacerda, 2011). Los eventos climáticos pueden influenciar en las especies parasitarias, en tanto que la antropización puede influenciar en la extinción de los parásitos sin afectar a sus huéspedes (de Melo et al., 2024).
Diversos estudios de la prevalencia de parasitismo reportan a las especies como Rhadinorhynchus sp., Camallanus sp., Lecithocladum sp. en otras especies comerciales como Scomberomorus commerson en Indonesia (Yusni et al., 2024) en tanto que, en tilapias provenientes de Etiopía se han reportado parásitos como Diplostomum sp., Contracaecum sp., Myxosporea sp. y Clinostomum spp. (Tesfay et al., 2024), estos hallazgos de prevalencias de diver-sidad de parásitos en especies acuícolas concuer-dan con un estudio realizado en la amazonía brasilera (estados de Amazonas y Acre) en el cual se reveló que la diversidad de especies parasitarias aumentó en ambientes conservados para hués-pedes detritívoros y omnívoros lo cual favorece el establecimiento de endoparasitos en base a nichos e infracomunidades (Virgilio et al., 2023).
Los nematodos presentan una enorme variedad de adaptaciones ecológicas (Roca, 1999) y junto con los trematodos son considerados los grupos taxonómicos más abundantes en peces de las riberas brasileñas (Virgilio et al., 2022). La mayoría son de vida libre y pueden encontrarse en el suelo, el agua dulce, el agua salobre y el océano. Otras son especies parásitas o semiparásitas que dañan tanto a plantas como a animales (Ringuelet, 1975; Chero et al., 2016). Alrededor del 40% de las 16,000 especies de nematodos que se han identificado son parásitos de animales. Solo el 8% de los nematodos parásitos conocidos infestan vertebrados, inclu-yendo a los humanos, y el resto infestan en invertebrados (Alvarado & Batanero, 2008).
El estudio de parasitismo en especies acuícolas amazónicas conlleva a realizar estudios posteriores en las cuales existe la posibilidad de seleccionar genotipos superiores para resistencia y resiliencia simultánea a parasitismo como la acantocefaliasis en el caso de la gamitana (Colossoma macropomum) (Agudelo et al., 2024).
El presente estudio evalúa la presencia de parásitos en P. squamosissimus procedentes de cuatro merca-dos de Iquitos, Loreto, Perú, evaluando la relación entre el peso y tamaño con mayor frecuencia de parásitos, con la finalidad de identificar la fauna parasitaria asociada, establecer una línea base y así asegurar la inocuidad de la especie comercializada a través de la creación de programas sanitarios y recomendaciones al público consumidor
METODOLOGÍA
Lugar de estudio
Los peces fueron obtenidos de cuatro mercados (“Modelo”, “Itinerante”, “Belén” y “Cardozo”) de la ciudad de Iquitos-Loreto, ubicadas en los distritos de Belén, Punchana y San Juan respectivamente. Estos son los lugares donde existe gran oferta y demanda de P. squamosissimus por parte de la población de la ciudad de Iquitos. La identificación de parásitos fue realizada en el Instituto de Investigación de la Amazonia Peruana (IIAP) en Iquitos y en el Laboratorio de Parasitología Animal de la Facultad de Medicina Veterinaria de la Universidad Peruana Cayetano Heredia (UPCH) en Lima.
Criterios de inclusión
Dentro de los criterios de inclusión se consideraron a ejemplares de P. squamosissimus con tamaño de entre 20 y 45 cm y peso de entre 300 y 600 g. Entre los criterios de exclusión a peces en estado de descomposición con mal olor o peces eviscerados.
Tamaño de muestra
La muestra estuvo constituida por 60 ejemplares de P. squamosissimus adquiridos de los mercados de estudio. Se tomó como referencia datos de estudios parasitológicos similares realizados en Brasil en P. squamosissimus en los cuales se empleó entre 44, 61 y 75 ejemplares de esta especie (Fontenelle et al., 2016; Chagas de Souza, 2020).
Procedimientos y técnicas
En la investigación se utilizó el muestreo probabilístico simple aleatorio, y los especímenes se eligieron directamente en los mercados para el respectivo análisis parasitológico, considerando siempre los criterios de inclusión y de exclusión de la muestra.
Obtención de muestras
Los especímenes fueron adquiridos durante los meses de diciembre (2021) a marzo (2022) en los mercados de Iquitos con una frecuencia de dos a tres veces por semana, a primera hora de la mañana, aproximadamente entre las 6:00 am y las 7:00 am. Basándose en la pigmentación de la piel y las branquias (color rojizo), además de intentar variar el tamaño, se eligieron los ejemplares que se encontraban en buen estado (frescos). Se tomaron medidas de longitud (total y estándar) y peso (en gramos) utilizando un ictiómetro graduado y una balanza, y los resultados se registraron en fichas de anotaciones. Posterior a la toma de datos biométricos se procedió al análisis parasitológico.
Análisis parasitológico
Procesamiento de nemátodos
Para la recolección de nemátodos se observó la cavidad celómica y mesenterio. Posteriormente el tracto digestivo fue abierto, colocado en una placa Petri y cubierto con agua destilada para su observación al estereoscopio (Figura 1). Con pinzas y estiletes se recogieron los nemátodos encon-trados. Se analizó el globo ocular, retirándolos y colocándolos en placas Petri para observación bajo estereoscopio.
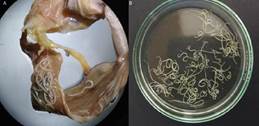
Figura 1. Larvas de Anisakidae colectadas de Plagioscion squamosissimus. A. Larvas adheridas al intestino de los peces. B. Larvas en placas Petri.
La musculatura fue analizada realizando cortes finos (fileteo de la carne). Los parásitos encontrados fueron colocados en placas Petri con solución salina al 0,85% para la limpieza con pinceles y estiletes. Finalmente, los parásitos fueron transferidos a viales o frascos de 10 ml conteniendo etanol al 70% para su conservación.
Los parásitos conservados fueron retirados de los frascos y colocados en láminas porta objeto con ácido láctico, cubiertos con láminas cubre objeto hasta la clarificación. Una vez clarificados los parásitos fueron observados bajo microscopio óptico para la identificación taxonómica.
Procesamiento de acantocéfalos
Se examinaron cuidadosamente muestras de intestino con el uso de estiletes y pinzas, teniendo cuidado de evitar que la probóscide se despren-diera y quedara en la pared intestinal. Los parásitos retirados de las muestras fueron colocados en placas Petri con agua destilada y refrigerados por 24 horas, con la finalidad de evaginar la probóscide. Para la fijación y conservación se utilizó AFA (95 partes de etanol 70 °GL, 3 partes de formalina (37% - 40%) y 2 partes de ácido acético glacial). Para la coloración de los acantocéfalos se utilizó el proceso regresivo de Carmín alcohólico de Langeron como descrito en la metodología de (Morey, 2019).
Identificación y caracterización de endoparásitos
Los parásitos se examinaron con un microscopio óptico LEICA DM750®; los parásitos se fotogra-fiaron utilizando una cámara digital acoplada al microscopio LEICA ICC50 La clasificación taxonó-mica se basó en la morfología y disposición de órganos internos de los parásitos, de acuerdo con Bush & Holmes (1986) y Moravec (1998).
La información recopilada fue procesada y anali-zada mediante estadística descriptiva. Se utilizó el coeficiente de correlación de Spearman (rs) para determinar la relación entre el tamaño del hospedador y la abundancia de parásitos. Se empleó el programa estadístico BioEstat 5,0 con un nivel de probabilidad menor de 0,05.
Análisis e interpretación de datos
Los índices parasitarios calculados fueron frecuen-cia, intensidad, intensidad media y abundancia media (Bush et al., 1997).
RESULTADOS Y DISCUSIÓN
Se identificaron siete especies de endoparásitos parasitando a P. squamosissimus: 6 pertenecientes a Nematoda: Anisakis larva tipo 1., Contracaecum sp. (larva), Hysterothylacium sp. (larva), Procamallanus (Spirocamallanus) inopinatus, Pseudoterranova sp. (larva), y Spiroxys sp. (larva) y una especie perteneciente a Acanthocephala: Rhadinorhynchus plagioscionis
Anisakis larva tipo 1
Orden: Ascaridida Skrjabin et Schulz, 1940
Familia : Anisakidae Railliet et Henry, 1912
Género: Anisakis Dujardin, 1945
Este nemátodo fue registrado en fase larval L3. Las principales características que permitieron su identificación taxonómica fueron: presencia de un mucron terminal en la parte posterior del cuerpo, un ventrículo prominente casi cuadrangular en la parte ventral del cuerpo y un diente larvario en la parte anterior del cuerpo (Figura 2).
Larva de Contracaecum sp.
Orden: Ascaridida Skrjabin et Schulz, 1940
Familia: Anisakidae Railliet et Henry, 1912
Género: Contracaecum Railliet et Henry, 1912
Este parásito fue registrado en fase larval L3. Las principales características que permiten su identificación son: presencia de diente larval, presencia de ventrículo pequeño de forma circular con orientación anterior del ciego intestinal ausencia de mucron en la cola (Figura 3).

Figura 2. Anisakis sp. A. Parte anterior mostrando diente larval. B. Parte media del cuerpo mostrando el ventrículo. C. Parte posterior mostrando el mucron.
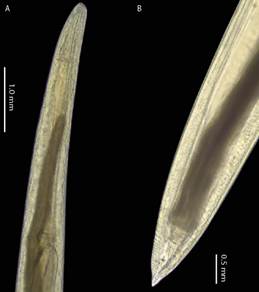
Figura 3. Contracaecum sp. A. Vistra ventral del cuerpo mostrando el ciego intestinal dirigido anteriormente. B. Parte posterior del cuerpo.
Larva de Hysterothylacium sp.
Orden: Ascaridida Skrjabin et Schulz, 1940
Familia: Anisakidae Railliet et Henry, 1912
Género: Hysterothylacium Ward et Magath, 1917
Este parásito se registró en fase larval L3. Las principales características de este parásito son: cuerpo alargado casi translúcido, terminación de la parte anterior en forma de punta roma, presencia de diente larval bastante pequeño, cola con terminación cónica.
Procamallanus (Spirocamallanus) inopinatus
Orden: Spirurida Chitwood, 1933
Familia: Camallanidae Railliet et Henry, 1915
Género: Procamallanus Baylis, 1923
Especie: Procamallanus (Spirocamallanus) inopinatus Travassos, Artigas et Pereira, 1928
Este parásito se identificó en fase adulta. Las principales características son cápsula bucal esclerotizada de color amarilla con anillos en espiral en número variable entre 18-22. Esófago muscular y glandular en forma de pera, siendo el glandular más grande que el muscular. Presencia de intestino desde el término del esófago glandular hasta la abertura anal. Hembra con terminación con proceso digitiforme. Macho con presencia de espículas y presencia de papilas genitales, siendo seis pre anales y cuatro posts anales (Figura 4).
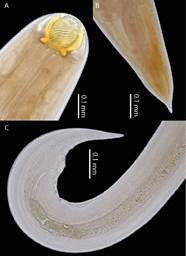
Figura 4. Procamallanus (Spirocamallanus) inopinatus. A. Parte anterior de ejemplar hembra mostrando la cápsula bucal. B. Parte posterior de ejemplar hembra. C. Parte posterior de ejemplar macho.
Larva de Pseudoterranova sp.
Orden: Ascaridida Skrjabin et Schulz, 1940
Familia: Anisakidae Railliet et Henry, 1912
Género: Pseudoterranova krabe, 1878
Fue identificado en fase larval L3. Es morfológica-mente similar a Anisakis larva tipo 1, presentando diente larval, mucrón y ventrículo prominente. La diferencia radica en la presencia de un ciego intestinal dirigido anteriormente que acompaña al ventrículo, ausente en Anisakis larva tipo 1 (Figura 5).

Figura 5. Pseudoterranova sp. A. Parte anterior mostrando el diente larval. B. Parte posterior mostrando el mucron. C. Vista ventral del cuerpo.
Spiroxys sp.
Orden: Spirurida Chitwood, 1933
Familia: Gnathostomatidae Railliet, 1895
Género: Spiroxys Schneider, 1866
Este nemátodo fue registrado en fase larval L3. Se caracteriza por tener la parte anterior en forma de “champiñón”, con dos pseudolabios esclerotizados en su parte media. El cuerpo es alargado, translúcido con terminación de la cola dirigida a un lado con punta redonda (Figura 6).

Figura 6. Spiroxis sp.
Rhadinorhynchus plagioscionis
Este acantocéfalo fue registrado en fase adulta. Tiene como característica principal la presencia de una probóscide bastante alargada cargada de espinas y ganchos.
De acuerdo con los resultados obtenidos en el presente estudio, P. squamosissimus posee una diversa fauna parasitaria de endoparásitos, lo cual coincide con varios estudios realizados en Brasil. En la Amazonía Brasileña, se han identificado géneros de nematodos como; Contracaecum, Terranova, Procamallanus, Thynnascaris y Anisakis (Rabelo et al., 2017; Fontenelle et al., 2016; Carva-lho et al., 2020); y los géneros de acantocéfalos Rhadinorhynchus y Neoechinorhynchus (Shagas de Souza, 2020). Anisakis larva tipo 1 registraron una frecuencia de parasitismo en la corvina del 56.6%, lo cual se aproxima a lo mencionado por Chagas de Souza et al. (2020) y Pereira et al. (2016) con prevalencias cercanas al 80%.
En cuanto al nemátodo Procamallanus (Spirocama-llanus) inopinatus se menciona que presenta baja especificidad de hospederos (Moravec, 1998). Ha sido reportada parasitando a más de 50 especies de peces de la Amazonía, principalmente de Brasil (Moravec, 1998). En el presente estudio, se reporta por primera vez parasitando a P. squamosissimus en la Amazonía peruana, aumentando su ocurren-cia en otro territorio correspondiente a la Amazonía.
Acantocéfalos de la Familia Rhadinorhynchidae presentan pocos géneros descritos: Rhadinor-hynchus; Polyacanthorhynchus y Pseudogorgo-rhynchus. Para P. squamosissimus fue descrita la especie Rhadinorhynchus plagioscionis en ejem-plares colectados en un mercado de la ciudad de Manaus, Amazonas-Brasil (Thatcher, 2006). En este estudio, esta especie de acantocéfalo fue registrada por primera vez en la Amazonía peruana, aumentando el rango de distribución geográfica de este parásito.
Asimismo, al encontrarse solo ejemplares larvales de Anisakis tipo 1 y Contracaecum sp. se considera a P. squamossisimus, como especie que participa en las interacciones depredador-presa en los ríos amazónicos y forma parte de la dieta de los mamíferos de agua dulce (Carvalho et al., 2020). Se deduce, que el primer hospedero intermediario de los nematodos serían los pequeños crustáceos y copépodos, lo cuales estarían presentes en los ríos amazónicos. El segundo hospedero intermediario serían los peces piscívoros como P. squamosissimus, finalmente las larvas infectivas pasarían a diversos hospederos definitivos de acuerdo a la especificidad del parasito.
El estudio más reciente para la Amazonía en donde se reportan especies zoonóticas en peces de consumo vendidos en los mercados de la ciudad, data del 2016. En este estudio, se reportó la presencia de Anisakis sp., Contracaecum sp., Terranova sp., Austrodiplostomum compactum; una especie de Denogeridae y una especie de acanto-céfalo: Rhadinorrhynchus plagioscionis (Pereira, 2016). En el presente estudio, se registran algunas de las mismas especies registradas en el 2016 en muestras procedentes de mercados de la ciudad de Manaus, Brasil.
Es posible que Anisakis spp. completen su ciclo vital en mamíferos acuáticos amazónicos, como ocurre con las especies marinas. Siendo el último hospedero algunas de las especies de delfines de agua dulce que son nativas de esta zona, incluyendo el delfín rosado (Inia geoffrensis) y el delfín negro (Sotalia fluviatilis) (Farias et al, 2017). Mientras que los hospederos finales del género Contra-caecum, Hysterothylacium y Pseudoterranova se presume serían las aves piscívoras, lagartos, peces carnívoros y mamíferos amazónicos respectiva-mente como lo mencionado por Moravec en 1998 y Klimpel & Palm en 2011. El poblador amazónico desempeñaría el rol del hospedero accidental a través del consumo de carne infectada de P. squamosissimus en preparaciones que empleen al pescado crudo o mal cocido.
En este estudio se registraron cuatro larvas de nemátodos con importancia en la salud pública alojados en órganos del tracto digestivo de P. squamosissimus, anisakídeos como: Anisakis larva tipo 1, Contracaecum sp., Pseudoterranova sp. e Hysterothylacium sp. lo cual concuerda con lo reportado por Farias et.al, (2017); Fontenelle et al, (2016) en Brasil. Dado que esta especie se comercializa en los mercados de Iquitos y que los pobladores amazónicos pueden preparar su carne de diversas formas, incluido el ceviche, el peligro de infección involuntaria por consumir carne de pescado parasitado existe y debe tenerse en cuenta antes de comerla.
Es crucial señalar que no todas las familias, géneros o especies de parásitos se consideran con potencial zoonótico, a pesar de que los peces de agua dulce tienen una ecología parasitaria diversa. La familia de nemátodos Anisakidae se destaca en esta categoría (Júnior & Da, 2005). En países como Japón, China, Corea, Holanda y Estados Unidos son frecuentes las infecciones zoonóticas, los proble-mas digestivos y respiratorios e incluso los casos de mortalidad (Jnior & Da, 2005).
En algunas ciudades brasileñas de Sudamérica se han documentado problemas zoonóticos relacio-nados con los anisakideos. (Ahmad et al., 2018) han sido atribuidos a la infección accidental de estos parásitos a través de la carne de pescado contaminada con los mismos (Ahmad et al., 2018; Carvalho et al., 2020; Fontenelle et al., 2016). En el Perú falta estudios y reportes de casos de anisakidosis en regiones amazónicas. Solo se tienen registros en el año 2004, en donde se describe ocho casos de la enfermedad en el país, todos ellos asociados al consumo de ceviche en zonas costeras (Lima e Ica).
A pesar de que la mayoría de los parásitos se encuentran en el aparato digestivo, algunas larvas de parásitos pueden atravesar la pared intestinal e introducirse en la musculatura. En varias especies zoonóticas de anisakideos, este comportamiento es típico. La ausencia de parásitos en la musculatura del pez no asegura que el consumidor de pescado no reciba carne contaminada con alguna toxina generada en algún momento por los parásitos, ya que los parásitos pueden emitir toxinas en el cuerpo del pez, y estas toxinas pueden acumularse en el músculo del pez (Morey, 2019). Los hallazgos del presente estudio son significativos para la salud pública porque muchas especies pueden suponer un riesgo al causar manifestaciones clínicas de enfermedades gastrointestinales, ya sea por su presencia física o por sustancias químicas secretadas. Es probable que muchos de estos síntomas estén presentes en los consumidores de carne de pescado todos los días, pero en ausencia de estudios especializados o de especialistas en la materia, pueden pasarse por alto o diagnosticarse erróneamente (Eiras et al., 2016; Sidney et al. 2020).
Índices parasitarios de P. squamosissimus pro-venientes de mercados de la ciudad de Iquitos, Perú
A todos los nemátodos detectados en el estudio se les determinaron índices parasitarios, dominando la larva de Anisakis tipo 1 con una frecuencia de 56,6%, intensidad de 537 individuos, intensidad media de 17,90 y abundancia media de 8,95. Los índices parasitarios más bajos los obtuvo Spiroxys sp., con frecuencias del 3,33%, intensidades de 2 individuos, intensidades medias de 1,00, y abun-dancias medias del 0,03% (Tabla 1).
Lugar de fijación de los parásitos presentes en P. squamosissimus
Los endoparásitos fueron registrados parasitando la cavidad visceral (458), intestino (421) y gónadas (30). De un total de 909 parásitos colectados, la mayoría provinieron de cavidad abdominal (458) e intestino (421). Estos resultados coinciden con lo observado en estudios realizados en Brasil, en donde se localizaron los parásitos en cavidad celómica, estomago e intestino de P. squamo-sissimus (Chagas de Souza, 2020; Seliz, 2022; Fontenelle et al., 2016; Lacerda et al., 2012; Eiras, 2010). Los peces albergan nemátodos adultos en sus tractos digestivos y cavidades corporales, por lo que la musculatura, el mesenterio, la cavidad corporal y los órganos internos puede albergar a una gran cantidad de individuos (Luque, 2004), representando un riesgo para los consumidores de pescado (Barros et al., 2006; Peña et al., 2013; Lerena et al., 2018).
La implicación practica del establecimiento de los perfiles más relevantes de poliparasitismo en P. squamosissimus, incluirían el conocimiento de la dinámica de las infestaciones asociadas con cada grupo de parásitos, lo cual permitiría entender las interacciones de un parasito con otro, así como en el medio ambiente y descubrir posibles causas de la presencia de ciertos grupos de nemátodos. Asimismo, se podrían precisar los perfiles de riesgo en la salud pública y planear intervenciones eficaces.
Asociaciones parasitarias
Se analizaron 60 ejemplares de P. squamosissimus, con rangos de peso de 300 a 600 gr y tamaño de 20 a 45 cm; 50 ejemplares estuvieron infectados por lo menos con una especie de parásitos. La frecuencia de monoparasitismo, fue 62% del total de los peces infectados.
Con respecto a los perfiles de poliparasitismo, se presentó biparasitismo en el 28% y triparasitismo en el 10% de los peces infectados. La diada más frecuente fue la producida por Anisakis larva tipo 1 y Pseudoterranova sp. representando el 64,3% del biparasitismo, seguida en un menor porcentaje de la producida por Anisakis larva tipo 1 y Procamallanus (Spirocamallanus) inopinatus con el 21,4%.
Tabla 1
Índices parasitarios de las especies de endoparásitos registrados en Plagioscion squamosissimus
|
Parásitos |
PA |
PP |
F% |
NTP |
IM |
AM |
|
Anisakis larva tipo 1 |
60 |
34 |
56,6 |
537 |
17,9 |
8,95 |
|
Contracaecum sp. (larva) |
60 |
6 |
10 |
6 |
1,00 |
0,10 |
|
Hysterothylacium sp. (larva) |
60 |
2 |
3,33 |
6 |
3,00 |
0,10 |
|
Procamallanus (spirocamallanus) inopinatus |
60 |
6 |
10,00 |
6 |
1,00 |
0,10 |
|
Pseudoterranova sp. (larva) |
60 |
15 |
25,00 |
327 |
21,8 |
5,45 |
|
Spiroxys sp. (larva) |
60 |
2 |
3,33 |
2 |
1,00 |
0,03 |
|
Rhadinorhynchus plagioscionis |
60 |
14 |
23,33 |
52 |
3,71 |
0,87 |
Leyenda: PA (Peces analizados), PP (Peces parasitados), F% (Frecuencia), NTP (Número total de parásitos), IM (Intensidad Media), AM (Abundancia Media).
La única triada de nematodos encontrados fue la compuesta por larvas de Anisakis larva tipo 1, Pseudoterranova sp. y Contracaecum sp. No se encontraron diferencias significativas en el número de parásitos por tamaño, peso y sexo (Tabla 2).
Tabla 2
Distribución de asociaciones parasitarias en Plagioscion squamosissimus de mercados de Iquitos, Perú
|
Parásito |
(+) |
Porcen- taje (%) |
|
Monoparasitismo |
|
|
|
Anisakis larva tipo 1 |
27 |
87,0 |
|
Procamallanus (Spirocamallanus) inopinatus |
2 |
6,5 |
|
Rhadinorhynchus plagioscionis |
2 |
6,5 |
|
SUBTOTAL |
31 |
62 |
|
Biparasitismo |
|
|
|
Hysterothylacium sp. (larva) + Spiroxys sp. (larva) |
2 |
14,3 |
|
Anisakis larva tipo 1 + Pseudoterranova sp.(larva) |
9 |
64,3 |
|
Anisakis larva tipo 1+ Procamallanus (Spirocamallanus) inopinatus |
3 |
21,4 |
|
SUBTOTAL |
14 |
28,0 |
|
Triparasitismo |
|
|
|
Anisakis larva tipo 1+ Pseudoterranova sp. (larva) + Contracaecum sp. (larva) |
5 |
100,0 |
|
SUBTOTAL |
5 |
10,0 |
|
TOTAL |
50 |
100% |
Con respecto a las asociaciones parasitarias encontradas, no existen estudios similares en peces de río con los cuales se pueda comparar. Se han hecho estudios para ver asociaciones en otros tipos de especies animales mas no en peces. En la presente investigación, la frecuencia de mono-parasitismo en P. squamosissimus, definida como la presencia de solo una especie de parasito infectando al hospedador, fue de 62%. Los casos con presencia de poliparasitismo, de dos o tres agentes, incluyeron larvas de los parásitos Anisakis tipo 1, Pseudoterranova sp., Procamallanus sp., Hysterohtylacium sp., Spiroxys sp. y Contracaecum sp., siendo la frecuencia de biparasitismo (28%) mayor a la del triparasitismo (10%). Chagas de Souza (2020) menciona de manera general, la presencia de por lo menos una especie de parasito infectando los ejemplares de P. squamosissimus, también refiere un predominio de ejemplares infectados por tres y cuatro especies de parásitos. Se encontraron varios perfiles de poliparasitismo intestinal, observándose en la mayoría de los especímenes el monoparasitismo y presentando un máximo de tres especies de parásitos por ejemplar en solo 5 peces. Mayores estudios permitirían tener una mejor estimación de la frecuencia, prevalencia y factores determinantes del poliparasitismo y así conocer el riesgo en la salud pública, además de priorizar y orientar recursos para su prevención y control.
Cálculo del Coeficiente de Correlación
El número total de endoparásitos detectados se correlaciono con el tamaño y el peso de los peces. Se obtuvo correlación positiva pero no estadística-mente significativa entre el tamaño y cantidad de endoparásitos; mientras que la correlación entre el peso y la cantidad de endoparásitos evidenció una correlación negativa, estadísticamente no signifi-cativa (rs = -0,04, p = 0,71; rs = -0,06, p = 0,59).
Uno de los factores por los que se encontraron estos parásitos con importancia en salud pública podría explicarse por su ciclo de vida complejo y heteroxeno, ya que requieren de una red alimen-taria de dos hospederos intermediarios para llegar al definitivo (Polley & Thompson, 2009). Asimismo, muchas especies de peces de consumo se consi-deran como hospederos intermediarios o paraté-nicos, lo cual puede incrementar el riesgo de infec-ción parasitaria con el tamaño del hospedador (Marcogliese, 2003). En el presente trabajo, sin embargo, no se registraron correlaciones estadís-ticamente significativas entre las variables en estudio, lo cual podría deberse al limitado rango de tamaño de peces adquiridos en la investigación.
CONCLUSIONES
A pesar del incremento de publicaciones relacio-nadas a la diversidad parasitaria en la Amazonía, muchos grupos de parásitos aún presentan incon-gruencias en su taxonomía debido a las limita-ciones en la identificación taxonómica tradicional. La presente investigación expande el rango de distribución geográfica de los nemátodos encon-trados y añade a P. squamosissimus dentro de la lista de hospederos paraténicos en Perú. Por ello se requiere el desarrollo de mayores estudios y la aplicación de herramientas moleculares para aclarar, con exactitud, la verdadera diversidad de parásitos que puedan afectar a los organismos acuáticos.
AGRADECIMIENTOS
Al Instituto de Investigaciones de la Amazonía Peruana (IIAP) por brindar las instalaciones del Laboratorio de Parasitología y Sanidad Acuícola para el desarrollo de la presente investigación.
REFERENCIAS BIBLIOGRÁFICAS
Agudelo, J. F., Neto, B. F., Mastrochirico-Filho, V. A., Ariede, R. B., Moreira, L. F., da Silva Noronha, L., & Hashimoto, D. T. (2024). Genetic parameters for host-response to acanthocephaliasis caused by the endoparasite Neoechinorhynchus buttnerae in the Amazon fish Colossoma macropomum. Aquaculture, 582, 740533.
Ahmad, F., Fazili, K. M., Sofi, O. M., Sheikh, B. A., & Sofi, T. A. (2018). Distribución patología causada por Bothriocephalus acheilognathi, Yamaguti 1934 (Cestoda: Bothriocephalidae). Revisión bibliográfica. Revista Veterinaria, 29(2), 142-149. https://doi.org/10.30972/vet.2923283
Alves, D. R., & Luque, J. L. (2001). Community ecology of the metazoan parasites of white croaker, Micropogonias furnieri (Osteichthyes: Sciaenidae), from the coastal zone of the State of Rio de Janeiro, Brazil. Memórias Do Instituto Oswaldo Cruz, 96, 145-153. https://doi.org/10.1590/S0074-02762001000200002
Alvarado, H., & Batanero, C. (2008). Significado del teorema central del límite en textos universitarios de probabilidad y estadística. Estudios pedagógicos (Valdivia), 34(2), 7-28.
Barros, L. A., Moraes, J., & Oliveira, R. L. de. (2006). Nematóides com potencial zoonótico em peixes com importância econômica provenientes do rio Cuiabá. Revista Brasileira de Ciência Veterinária, 13(1), 55-57. https://doi.org/10.4322/rbcv.2014.267
Brack, A. (1997). Pobreza y manejo adecuado de los recursos en la Amazonía peruana. Revista Andina, 29, 15-39.
Bush, A. O., & Holmes, J. C. (1986). Intestinal helminths of lesser scaup ducks: An interactive community. Canadian Journal of Zoology, 64(1), 142-152. https://doi.org/10.1139/z86-023
Bush, A. O., Lafferty, K. D., Lotz, J. M., & Shostak, A. W. (1997). Parasitology meets ecology on its own terms: Margolis et al. revisited. The Journal of Parasitology, 83(4), 575-583. https://doi.org/10.2307/3284227
Carvalho, E. L. de, Santana, R. L. S., Gonçalves, E. C., Pinheiro, R. H. da S., & Giese, E. G. (2020). Primeiro relato de Anisakis sp. (Nematoda: Anisakidae) parasitando pato doméstico na Ilha de Marajó, estado do Pará, Brasil. Revista Brasileira de Parasitologia Veterinária, 29. https://doi.org/10.1590/S1984-29612020015
Chagas de Souza, D., Eiras, J. C., Adriano, E. A., & Corrêa, L. L. (2020). Metazoan parasites of Plagioscion squamosissimus (Osteichthyes: Sciaenidae) of two rivers from the eastern Amazon (Brazil). Annals of Parasitology, 66(2), 217-225. https://doi.org/10.17420/ap6602.257
Chappel, L. (1994). Diplostomiasis: The disease and host-parasite interactions. Parasitic diseases of fish. https://ci.nii.ac.jp/naid/10019344052
Chero, J., Sáez, G., Iannacone, J., Cruces, C., Alvariño, L., & Luque, J. (2016). Ecología comunitaria de metazoos parásitos del bonito Sarda chiliensis Cuvier, 1832 (Perciformes: Scombridae) de la costa peruana. Revista de Investigaciones Veterinarias del Perú, 27(3), 539-555. https://doi.org/10.15381/rivep.v27i3.12008
de Melo, H.P.S., Takemoto, R.M., Gonçalves, G.S.R. et al. (2024). Effects of climate change on the distribution of Hoplias malabaricus and its ecto- and endoparasite species in South America. Aquat Ecol 58, 999–1011. https://doi.org/10.1007/s10452-024-10120-7
Eiras, J., Takemoto, R., & Pavanelli, G. C. (2010). Diversidade de parasitas de peixes de água doce do Brasil.
Eiras, J. C., Pavanelli, G. C., Takemoto, R. M., Yamaguchi, M. U., Karkling, L. C., & Nawa, Y. (2016). Potential risk of fish-borne nematode infections in humans in Brazil – Current status based on a literature review. Food and Waterborne Parasitology, 5, 1-6. https://doi.org/10.1016/j.fawpar.2016.08.002
Farias Rabelo, N. L., Muniz e Silva, T. C., Ferreira Araujo, L. R., da Silva Pinheiro, R. H., & Machado da Rocha, C. A. (2017). Detection of Anisakidae larvae parasitizing Plagioscion squamosissimus and Pellona castelnaeana in the State of Pará, Brazil. Acta Scientiarum. Biological Sciences, 39(3), 389-395.
Fontenelle, G., Knoff, M., Felizardo, N. N., Torres, E. J. L., Matos, E. R., Gomes, D. C., & São Clemente, S. C. de. (2016). Anisakid larva parasitizing Plagioscion squamosissimus in Marajó Bay and Tapajós River, state of Pará, Brazil. Revista Brasileira de Parasitologia Veterinária, 25, 492-496. https://doi.org/10.1590/S1984-29612016034
Galvis, G., Mojica, J., Duque, S., Castellanos, C., Sánchez Duarte, P., Arce H., M., Gutiérrez-Cortés, A., Jiménez-Segura, L., Santos-Acevedo, M., Rivadeneira, S., Arbeláez, F., Prieto Piraquive, E., & Leiva, M. (2006). Peces del medio Amazonas.
García Dávila, C., Sánchez Riveiro, H., Flores Silva, M. A., Mejía de Loayza, E., Angulo Chávez, C., Castro Ruiz, D., Estivals, G., García Vásquez, A., Nolorbe Payahua, C., Vargas Dávila, G., Núñez, J., Mariac, C., Duponchelle, F., & Renno, J.-F. (2018). Peces de consumo de la Amazonía peruana. Instituto de Investigaciones de la Amazonía Peruana. http://repositorio.iiap.gob.pe/handle/20.500.12921/369
García-Vásquez, A., Vargas, G., Tello-Martín, S., & Duponchelle, F. (2012). Desembarque de pescado fresco en la ciudad de Iquitos, región Loreto - Amazonía peruana. Folia Amazónica, 21(1-2), 45-52. https://doi.org/10.24841/fa.v21i1-2.31
Júnior, S., & Da, E. A. (2005). Manual de controle higiênico-sanitário em serviços de alimentação (p. 623).
Lacerda, A. C. F., Takemoto, R. M., Tavares-Dias, M., Poulin, R., & Pavanelli, G. C. (2012). Comparative parasitism of the fish Plagioscion squamosissimus. The Journal of Parasitology, 98, 713-717.
Lacerda, A. C. F. (2011). Endoparásitos de tucunaré-azul (Cichla piquiti, Cichlidae) y corvina (Plagioscion squamosissimus, Sciaenidae) en bacias hidrográficas nativas e invadidas: Testando a hipótese do escape do inimigo. https://aquadocs.org/handle/1834/10074
Lerena, M. S. M., Torres, E. P., Ramírez, M. Á. P., Orozco, J. A. H., Estrada, G. E., & Esparza, C. D. E. (2018). Presencia de parásitos en pescado en el mercado local de la Zona Metropolitana de Guadalajara. e-CUCBA, 10, 21-26. https://doi.org/10.32870/e-cucba.v0i10.117
Lizama, M. de L. A. P., Takemoto, R. M., & Pavanelli, G. C. (2006). Parasitism influence on the hepato, splenosomatic and weight/length relation and relative condition factor of Prochilodus lineatus (Valenciennes, 1836) (Prochilodontidae) of the Upper Paraná River floodplain, Brazil. Revista Brasileira de Parasitologia Veterinária = Brazilian Journal of Veterinary Parasitology, 15(3), 116-122.
Luque, J. (2004). Biologia, epidemiologia e controle de parasitos de peixes. XIII Congresso Brasileiro de Parasitologia Veterinária & I Simpósio Latino-Americano de Ricketisioses, Ouro Preto, MG.
Machado, P. M., Takemoto, R. M., & Pavanelli, G. C. (2005). Diplostomum (Austrodiplostomum) compactum (Lutz, 1928) (Platyhelminthes, Digenea) metacercariae in fish from the floodplain of the Upper Paraná River, Brazil. Parasitology Research, 97(6), 436-444. https://doi.org/10.1007/s00436-005-1483-7
Matta, R. (2014). República gastronómica y país de cocineros: Comida, política, medios y una nueva idea de nación para el Perú. Revista Colombiana de Antropología, 50(2), 9-13.
Marcogliese, D. J. (2003). Food webs and biodiversity: Are parasites the missing link? Journal of Parasitology, 89(6), 106–113.
Ministerio de la Producción (PRODUCE). (2019). Anuario estadís-tico pesquero y acuícola 2019.
Moravec, F. (1998). Nematodes of freshwater fishes of the Neotropical region. Institute of Parasitology, Academy of Sciences of the Czech Republic.
Murrieta Morey, G. (2019). Parasitología en peces de la Amazonía: Fundamentos y técnicas parasitológicas, profilaxis, diagnóstico y tratamiento. Instituto de Investigaciones de la Amazonía Peruana. http://repositorio.iiap.gob.pe/handle/20.500.12921/394
Nachev, M., Rozdina, D., Michler-Kozma, D. N., Raikova, G., Sures, B. (2022). Metal accumulation in ecto- and endoparasites from the anadromous fish, the Pontic shad (Alosa immaculata). Parasitology, 149(4), 496-502. https://doi.org/10.1017/S0031182021002080
Peña, Y. P., Castillo, V. L., & Almanza, D. Á. (2013). Parásitos de transmisión alimentaria en Cuba: Una revisión de la literatura. Revista Cubana de Alimentación y Nutrición, 23(1), 130-138.
Pereira, N. R. B. (2016). As espécies parasitas com potencial zoonótico em peixes amazônicos. 152-152.
Polley, L., & Thompson, R. C. (2009). Parasite zoonoses and climate change: Molecular tools for tracking shifting boundaries. Trends in Parasitology, 25(6), 285–291.
Ranzani-Paiva, M. J. T., Silva-Souza, A. T., Pavanelli, G. C., & Takemoto, R. M. (2000). Hematological characteristics and relative condition factor (Kn) associated with parasitism in Schizodon borellii (Osteichthyes, Anostomidae) and Prochilodus lineatus (Osteichthyes, Prochilodontidae) from Paraná river, Porto Rico region, Paraná, Brazil. Acta Scientiarum, 22(2), 515-521.
Ricker, W. (1975). Computation and interpretation of biological statistics of fish populations. Bulletin of the Fisheries Research Board of Canada, 191, 1-382.
Ringuelet, R. A. (1975). Zoogeografía y ecología de los peces de aguas continentales de la Argentina y consideraciones sobre las áreas ictiológicas de América del Sur. Ecosur, 2(3).
Roca, V. (1999). Relación entre las faunas endoparásitas de reptiles y su tipo de alimentación. 21.
Serrano-Martínez, E., H, M. Q., M, E. H., & P, L. P. (2017). Detección de parásitos en peces marinos destinados al consumo humano en Lima Metropolitana. Revista de Investigaciones Veterinarias del Perú, 28(1), 160-168.
Sidney, M., Oliveira, B., Lima Corrêa, L., Drielly, O., & Ferreira, M. (2020). Larvae of zoonotic potential nematodes infecting carnivorous fish from the lower Jari River in Northern Brazil. Biota Amazônia, 9(4), 50-52. https://doi.org/10.18561/2179-5746/biotaamazonia.v9n4p50-52
Takemoto, R. M., Pavanelli, G. C., Lizama, M. A. P., Lacerda, A. C. F., Yamada, F. H., Moreira, L. H. A., Ceschini, T. L., & Bellay, S. (2009). Diversity of parasites of fish from the Upper Paraná River floodplain, Brazil. Brazilian Journal of Biology, 69, 691-705. https://doi.org/10.1590/S1519-69842009000300023
Tello, H., & Alcántara, F. (2007). Potencial amazónico: Acerca del desarrollo de la pesquería y piscicultura en la Amazonía peruana. Perú Económico, 30(6).
Tesfay, S., Teferi, M., Dejenie, T., Abay, T., & Hiluf, G. (2024). A survey of ecto and endoparasites of Nile tilapia, Oreochromis niloticus (Linnaeus, 1758) fingerlings in Midmar reservoir, Adwa, northern Ethiopia. Journal of Applied Animal Research, 52(1). https://doi.org/10.1080/09712119.2024.2310158
Thatcher, V. E. (2006). Amazon fish parasites. In J. Adis, J. R. Arias, G. Rueda-Delgado, & K. M. Wantzen (Eds.), Aquatic biodiversity in Latin America (pp. 508). Sofia: Pensoft.
Virgilio, L. R., Martins, W. M. O., Lima, F. S, Takemoto, R. M., Camargo, L. M. A., & Meneguetti, D. U. O. (2022). Endoparasite fauna of freshwater fish from the upper Juruá River in the Western Amazon, Brazil. Journal of Helminthology, 96, e55.
Virgilio, L.R., de Melo, H.P.S., da Silva Lima, F. et al. (2023). Fish endoparasite metacommunity in environments with different degrees of conservation in the western Brazilian Amazon. Parasitol Res, 122, 2773–2793 https://doi.org/10.1007/s00436-023-08013-z
Yusni, E., Batubara, A., & Frantika, C. (2022). Detection of endoparasites in mackerel tuna (Euthynnus affinis) in north Sumatra province, Indonesia. Iraqi Journal of Veterinary Sciences, 36(2), 519-524.