INTRODUCCIÓN
Native edible mushroom "Qipatari" (Basidiomycota): Strain production, mycelial growth on lignocellulosic substrates, fruiting body production, and nutritional content
Karin Antonieta Pérez-Leguía1, 3, 4 *; Albino Quispe-Pelaez1, 2; María Encarnación Holgado-Rojas1, 3
1 Centro de Investigación y Producción de Hongos Alimenticios y Medicinales (CIPHAM), Facultad de Ciencias Biológicas, Universidad Nacional de San Antonio Aban del Cusco. Perú.
2 Escuela Profesional de Ingeniería Agropecuaria- Filial Andahuaylas, Facultad de Agronomía y Zootecnia. Universidad Nacional de San Antonio Aban del Cusco. Perú.
3 Organización para la Mujer en la Ciencia para el Mundo en Desarrollo (OWSD).
4 Departamento Académico de Ciencias Básicas, Universidad Nacional José María Arguedas, Andahuaylas, Perú.
* Autor corresponsal: karin.perez@unsaac.edu.pe (K. A. Pérez-Leguía).
ORCID de los autores:
K. A. Pérez-Leguía: https://orcid.org/0000-0002-1712-1402 A. Quispe Pelaez: https://orcid.org/0000-0002-2436-1455
M. E. Holgado-Rojas: https://orcid.org/0000-0002-2285-8679
RESUMEN
Los hongos comestibles nativos conctituyen una fuente potencial de alimento y compuestos bioactivos con aplicaciones nutracéuticas. En este estudio, se evaluó el cultivo y el valor nutricional del hongo nativo "Qipatari", recolectado en la localidad de Río Blanco (Distrito Los Chankas), Apurímac, Perú. Los objetivos también incluyeron la obtención de una cepa del hongo, su cultivo controlado y la determinación del contenido nutricional de los carpóforos tanto silvestres como cultivados. El análisis molecular preliminar del hongo mostró similitudes del 88,30% - 89,97% con Lentinus levis y del 88,49% con Pleurotus cytidiosus var. formosensis. La fructificación fue óptima en un sustrato compuesto por 80% de rastrojo de trigo y 20% de aserrín de aliso. Se identificaron ocho aminoácidos esenciales, destacando la alanina (268,1 mg/100 g) y la lisina (123,0 mg/100 g), además de ácidos grasos como el ácido linoleico (3,288 mg/100 g) y azúcares libres como fructosa y glucosa. Los carpóforos silvestres y cultivados presentaron un mayor contenido de ceniza, fibra y calcio en comparación a los hongos comerciales Lentinula edodes y P. ostreatus, mientras que el contenido proteico fue similar. El reporte de la comestibilidad del “Qipatari”, su cultivo y contenido nutricional, son nuevos aportes para la ciencia, proporcionando una base para su producción comercial y conservación.
Palabras clave: nutracéutico, hongo nativo; Río Pampas; Apurímac; bosque mesoandino.
ABSTRACT
Native edible mushrooms represent a potential source of food and bioactive compounds with nutraceutical applications. In this study, the cultivation and nutritional value of the native mushroom "Qipatari," collected in the town of Rio Blanco (Los Chankas District), Apurimac, Peru, were evaluated. The objectives were to obtain the mushroom strain, perform its cultivation, and determine the nutritional content of wild and cultivated carpophores. Preliminary molecular analysis of the fungus showed similarities of 88.30% – 89.97% with Lentinus levis and 88.49% with Pleurotus cytidiosus var. formosensis. Fruiting was optimal in a substrate composed of 80% wheat stover and 20% alder sawdust. Eight essential amino acids were identified, most notably alanine (268.1 mg/100 g) and lysine (123.0 mg/100 g), in addition to fatty acids such as linoleic acid (3.288 mg/100 g) and free sugars such as fructose and glucose. Wild and cultivated carpophores had higher ash, fiber, and calcium contents than commercial mushrooms Lentinula edodes and P. ostreatus, while protein content was similar. The reported edibility of “Qipatari”, its cultivation, and nutritional content are new contributions to science, providing a basis for its commercial production and conservation.
Keywords: nutraceutical; native fungus; Pampa River; Apurimac; mesoandean forest.
Recibido: 20-01-2025.
Aceptado: 10-05-2025
INTRODUCCIÓN
El conocimiento micológico tradicional es considerado un legado ancestral que forma parte de la identidad biocultural de las comunidades. Este conocimiento no solo incluye la identificación de especies, sino también prácticas relacionadas con su recolección y preparación (Dávila-Arenas et al., 2013, Ramírez-Terrazo et al., 2021), esta es la base para el aprovechamiento de los hongos silvestres, pues permite identificar y conocer las especies que pueden ser potencialmente susceptibles de cultivo, con fines de consumo y comercialización, principalmente aquellas que son valoradas regionalmente. El estudio de especies naturales de hongos comestibles, podría ser una práctica útil para mejorar los rendimientos, introducir nuevas especies en los mercados y preservar el germoplasma de especies de hongos antes de que sus entornos naturales sean alterados o destruidos por la acción humana, ya que estas cepas respaldarán la futura producción mundial de hongos. Así mismo, la fructificación de los hongos depende de las condiciones climáticas, por lo que los rendimientos anuales en la naturaleza pueden cambiar de un año a otro, dependiendo de variables como la temperatura media o la tasa de precipitación; por lo tanto, es predecible y ya está demostrado que la domesticación de algunas especies de hongos proporciona una productividad estable independientemente del cambio climático (Lallawmsanga & Carrasco, 2022).
Sin embargo, Waiganjo et al. (2023) destacan que, a pesar del creciente interés en los hongos comestibles por sus beneficios nutricionales y económicos, muchas especies silvestres aún no se cultivan ampliamente debido a barreras técnicas y económicas relacionadas con su domesticación y comercialización sostenible, sumado a que, algunas especies son consideradas no cultivables (Cano & Romero, 2016) ya sea por factores biológicos intrínsecos, o por sus requerimientos nutricionales específicos tanto en la formulación de los ingredientes como en el proceso de cultivo (Zied et al, 2024).
La necesidad de explorar y documentar estas especies es crucial, tomando en cuenta el vasto número de hongos que son utilizados como alimento, medicina o ambos, para la supervivencia, las culturas y las economías de cientos de grupos étnicos en todo el mundo (Pérez-Moreno, 2021). Así mismo su cultivo y producción permiten valorar los subproductos lignocelulósicos generados en la silvicultura y la agricultura, siendo un modelo de agricultura circular rentable y carbono-positivo con menores emisiones de gases de efecto invernadero (GEI) que otros sectores, contribuyendo a la sostenibilidad de los bosques y el medio ambiente circundante (Jasinska, 2023; Okuda, 2022). Ello cobra mayor relevancia dado que actualmente la quema de residuos de cultivos en el campo abierto se ha convertido en una preocupación importante para los esfuerzos de mitigación del cambio climático en todo el mundo, requiriéndose la adopción de prácticas de gestión de residuos sostenibles que respeten y salva-guarden el medio ambiente y la salud de las personas (Raza et al., 2022).
El uso de hongos comestibles en América se remonta a las culturas precolombinas (Pérez-Moreno, 2021), y en regiones andinas y selváticas del Perú, el conocimiento etnomicológico sigue siendo de gran relevancia (Pavlich, 2001; Chimey & Holgado, 2010; Trutmann et al., 2012). Sin embargo, en la región Apurímac, aún no se han realizado estudios etnomicológicos o de cultivo, por lo que el presente estudio se propone llenar este vacío científico al ser el primer reporte sobre el hongo comestible conocido como "Qipatari", que crece en los bosques nativos mesoandinos del valle del río Pampas (Provincia de Chincheros, Perú) siendo recolectado y consumido por la población local desde tiempos antiguos.
El objetivo principal del presente estudio es aislar la cepa del hongo, determinar la formulación óptima de sustratos lignocelulósicos para su cultivo, producción de cuerpos fructíferos y analizar su contenido nutricional. Con ello, se busca no solo contribuir a la diversificación de la oferta alimentaria resiliente frente al cambio climático, sino también incentivar la valorización, conserva-ción y manejo sostenible de los bosques nativos que aun albergan una biodiversidad, especialmente micológica, en gran parte desconocida.
METODOLOGÍA
Debido a que el “Qipatari” es un hongo silvestre, sin antecedentes de cultivo, se procuró seguir las recomendaciones y pasos para la domesticación de especies naturales de hongos comestibles mencionada por Albertó (2017), con ciertas modificaciones (Figura 1).
Colecta
Se realizó la búsqueda intensiva de los carpóforos del “Qipatari” durante la época lluviosa (mes de marzo del 2018) en los bosques mesoandinos nativos del distrito Los Chankas, Provincia de Chincheros, Departamento de Apurímac. Se colectaron los basidiocarpos que presentaron las mejores características en cuanto al tamaño, aspecto y textura, los cuales fueron extraídos de su hábitat con la ayuda de una navaja (Aguilar-Pumahuillca et al., 2019) y colocados en recipientes plásticos estériles y herméticos, para ser transportados hasta el Centro de Investigación y Producción de Hongos Alimenticios y Medicinales (CIPHAM) de la Facultad de Ciencias Biológicas de la Universidad Nacional de San Antonio Abad del Cusco, Provincia y Departamento del Cusco.
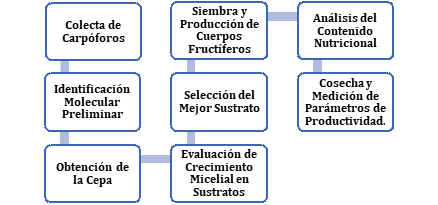
Figura 1. Flujograma metodológico del estudio.
Identificación molecular preliminar
Este procedimiento, en su totalidad, fue realizado por el Departamento de Filogenética y Evolución Molecular del Centro de Investigación Biológica y Química de la Universidad de Varsovia (Polonia), donde se enviaron muestras secas de los carpóforos, para extracción, amplificación y secuenciación de ADN, para lo cual, emplearon los oligonucleótidos universales para hongos: ITS1 e ITS4. Las secuencias obtenidas fueron comparadas con las depositadas en el GenBank mediante el software BLAST, calculando el porcentaje de similitud con las especies más cercanas.
Obtención de la cepa
Se seleccionaron los carpóforos más idóneos considerando el tamaño y el grado de desarrollo, preferentemente los juveniles con buenas características organolépticas, luego en una cámara de flujo laminar, se realizaron cortes de 0.4-0.5 cm del pseudotejido y se colocaron en 6 cajas petri (con 2 tipos diferentes de medio de cultivo: 3 con Papa Dextrosa Agar (PDA) y 3 con Extracto de Malta Agar (EMA) (Arana-Gabriel, 2020). Todas fueron incubadas a 25 °C durante dos semanas (Incubadora microbiológica MEMMERT (LP-111), Alemania). Se llevaron a cabo revisiones constantes del crecimiento micelial y se descartaron las placas contaminadas. Posteriormente, se realizaron los repiques necesarios para purificar la cepa, hasta obtener una cepa axénica, libre de contaminantes, que se incorporó en el cepario del CIPHAM.
Evaluación del crecimiento micelial lineal en sustratos lignocelulósicos
Se seleccionaron residuos lignocelulósicos teniendo en cuenta el sustrato donde se había encontrado el hongo y su disponibilidad en el entorno. Dada la falta de antecedentes sobre su cultivo, se desarrollaron cinco fórmulas basadas en estudios previos que han demostrado la eficacia de diversos residuos lignocelulósicos como sustratos para el cultivo de hongos (Holgado et al., 2019; Albertó, 2008; Mushworld, 2005). Este enfoque no solo buscó optimizar las condiciones para el cultivo del hongo "Qipatari", sino también contribuir a la sostenibilidad mediante el uso de residuos agrícolas predominantes, promoviendo así un manejo más responsable y eficiente de los recursos disponibles. Las formulaciones evaluadas (F), fueron: F1= 80% Aserrín de Aliso + 20% Rastrojo de trigo; F2=80% Aserrín de eucalipto y 20% rastrojo de trigo; F3= 80% rastrojo + 20% aserrín de aliso; F4=80% rastrojo y 20% de Aserrín de eucalipto; F5=Chala de maíz el 100%; finalmente se añadió 1% de yeso en función del peso seco de cada formulación, y se hidrató cada mezcla hasta alcanzar un 65% de humedad (Anexo 1).
Para determinar el sustrato óptimo, se empleó la prueba de crecimiento micelial lineal (Curvetto et al., 2002) con algunas modificaciones, empleán-dose tubos de ensayo de 9 cm de longitud y 1,6 cm de diámetro, donde se colocó cada formulación con cinco repeticiones, esterilizándose a 121°C y 2 Bares de presión (Autoclave WOLF (KL-71), Argentina). Todos los tubos, una vez enfriados, fueron inoculados con un disco de micelio de 1 cm, utilizando una Cámara de flujo laminar (BioBase, China). Los extremos de los tubos se cubrieron con tapones de algodón para ser incubados a 25 °C. El desarrollo micelial fue medido diariamente hasta alcanzar una longitud de 7 cm. Los datos obtenidos fueron analizados mediante un ANVA (Análisis de Varianza) y Prueba de Tukey, lo que permitió seleccionar las formulaciones de los subgrupos con las mejores medias y más semejantes entre sí para la siembra final. Esta metodología asegura la elección de los sustratos más adecuados para ser empleados en la siguiente etapa experimental.
Siembra y producción de cuerpos fructíferos
Inicialmente, para producir la "semilla fúngica o spawn", se usaron granos de trigo (Triticum aestivum) como base, siguiendo el método descrito por Yamile & Fabio (2001). La inoculación se llevó a cabo en una cámara de flujo laminar, utilizando improntas de micelio de 2,5 cm. Las bolsas inoculadas se incubaron a 25 °C en completa oscuridad, conforme a lo mencionado por López (2007) hasta que el micelio cubriera todos los granos.
Las formulaciones con los mejores resultados del experimento anterior se hidrataron durante un día, añadiendo agua destilada hasta alcanzar un 65% de humedad. Posteriormente se incorporó yeso en una proporción del 1% del peso seco total de cada formulación para lograr un pH óptimo del sustrato (Albertó 2008). Finalmente, los sustratos se colocaron en bolsas de polipropileno de 500 g (Mushworld, 2005) y se sometieron a esterilización por autoclave durante 2 horas a 121ºC y 2 Bares de presión. Luego se dejaron enfriar a temperatura ambiente, lo cual es crucial para evitar daños en el sustrato y asegurar que se mantenga en condiciones óptimas para la inoculación. Seguidamente, en una cámara de flujo laminar se procedió a realizar la siembra inoculando 50g de semilla por bolsa, que representa una Tasa de inoculación del 10% (Sánchez & Royse 2001). La incubación se llevó a cabo a 25 °C y en total oscuridad para evitar la inhibición del crecimiento del micelio (Chang & Miles, 1989), este proceso se mantuvo por 60 días, momento en el cual se observó la completa colonización del micelio en el sustrato, y que éste mantuviera su consistencia al retirar la bolsa (Omarini et al., 2009).
Las condiciones controladas para el cultivo del hongo “Qipatari” se establecieron en un ambiente específico, donde se mantuvieron parámetros óptimos de temperatura y humedad. A continua-ción, se detallan las prácticas y condiciones implementadas:
La temperatura se mantuvo a 13 °C con una variación de ±1°C. Este rango es adecuado para el crecimiento de hongos, ya que temperaturas entre 10 °C y 25 °C son generalmente óptimas para diversas especies de hongos
La Humedad Relativa se controló al 90% con una tolerancia de ±5%. Este nivel de humedad es crítico para maximizar el crecimiento, ya que la mayoría de los cultivos en invernadero requieren una humedad relativa alrededor del 80% para un desarrollo óptimo.
Inicialmente, se realizaron riegos dos veces al día utilizando un pulverizador manual. Una vez que aparecieron los primordios del hongo, la frecuencia de riego se redujo a una vez al día. Este ajuste es común en el cultivo de hongos, donde la humedad debe ser alta durante las etapas iniciales y puede disminuir a medida que avanza el desarrollo.
Se garantizó una ventilación adecuada durante 2 horas diarias mediante un ventilador con filtro conectado a una ventana exterior. La ventilación es esencial para evitar la acumulación de CO₂ y mantener un ambiente saludable para los hongos
La iluminación se optimizó con lámparas fluorescentes encendidas entre 10 y 12 horas diarias. La luz juega un papel importante en la inducción del crecimiento de ciertos hongos, aunque muchos no requieren luz directa para su desarrollo.
Pruebas piloto
Antes de estandarizar estos parámetros, se llevaron a cabo pruebas piloto que permitieron identificar los valores óptimos necesarios para inducir la fructificación del hongo “Qipatari”. Estos ensayos fueron fundamentales para adaptar las condiciones a las necesidades específicas del hongo y maximizar su crecimiento y desarrollo.
Una vez que los cuerpos fructíferos emergieron y alcanzaron un desarrollo adecuado, se llevó a cabo la cosecha de los carpóforos. Este proceso incluyó varias etapas importantes:
Cosecha manual: Los carpóforos se cortaron cuidadosamente con una cuchilla limpia y desinfectada, asegurándose de no dejar fragmentos que pudieran contaminar el cultivo. Esto es esencial para prevenir la proliferación de enfermedades en el sustrato.
Pesaje: Los ejemplares cosechados fueron pesados inmediatamente en estado fresco. Este pesaje se registró de acuerdo con la numeración de las bolsas y la formulación del sustrato de donde se recolectaron.
Mediciones del Píleo: Se tomaron medidas del píleo, que es la parte superior del hongo, lo que permite evaluar su tamaño y calidad. Estas mediciones son relevantes para determinar las características del producto cosechado y su idoneidad para el consumo y/o comercialización (Staments & Chilton, 1993; Femández, 2005).
Cálculo de Parámetros de productividad: Con los datos recopilados, se calcularon los parámetros de productividad, permitiendo así evaluar el rendimiento del cultivo en función de las diferentes formulaciones utilizadas. Esto ayuda a identificar qué condiciones o sustratos son más efectivos para maximizar la producción del hongo. La eficiencia biológica (EB) se halló a partir del peso fresco total de cuerpos fructíferos (g)/peso seco inicial del sustrato x 100 (Naim et al., 2020). El rendimiento se representó como la relación en porcentaje entre el peso fresco del hongo y el peso del sustrato húmedo (De Jesús-Rivera et al., 2022). La tasa de productividad se calculó dividiendo la eficiencia biológica en cada sustrato y el tiempo transcurrido en días desde la incubación hasta la cosecha (Tekeste et al., 2020).
Este enfoque sistemático no solo asegura una cosecha eficiente, sino que también proporciona información valiosa para futuras siembras y mejoras en las prácticas de cultivo.
Análisis del contenido nutricional
Los carpóforos silvestres y cultivados se dejaron secar al ambiente durante tres días. Posterior-mente, se empaquetaron en bolsas de polipro-pileno para ser transportados al Laboratorio de Análisis Fisicoquímicos y al Laboratorio de Análisis Cromatográficos, quienes se encargaron de la evaluación de proteínas, carbohidratos, grasas, minerales, azúcares, aminoácidos, y ácidos grasos. Ambos laboratorios son parte de la Unidad de Prestaciones de Servicios de la Escuela Profesional de Química de la UNSAAC.
Evaluación estadística
Se empleó el diseño experimental completamente al azar (DCA) con un nivel de significancia del 5% (p < 0,05) y el Análisis de Varianza (ANOVA) para comparar las medias de crecimiento micelial entre las diferentes formulaciones de sustrato. Para determinar diferencias significativas entre los tratamientos, se utilizó la prueba de Tukey. Los datos obtenidos se analizaron utilizando el software RStudio 2024 (versión 12.0).
RESULTADOS Y DISCUSIÓN
Los carpóforos del hongo comestible “Qipatari”, fueron recolectados en la ubicación UTM 632811 18L y 8514882, de la localidad de Río Blanco (Distrito Los Chankas, Apurímac) (Anexo 2) a una altitud de 2884 metros. El hongo (Figura 2), de hábito lignocelulósico, se halló creciendo sobre troncos muertos de Alnus acuminata ("lambras") y Myrcianthes oreophila ("unka").
Durante la recolección, se obtuvieron 4 ejemplares, de estos, 3 se encontraban en buen estado y 01 se encontró en la última etapa de su desarrollo.
El nombre “Qipatari” (palabra del quechua chanca), que quiere decir “el que se encuentra después” (traducción basada en Zariquiey & Córdova (2008)), hace referencia a la aparición tardía de este hongo en los bosques mesoandinos, que, según los pobladores, suele hallarse bien entrada la época lluviosa. Este proceso de recolección es signifi-cativo no solo por la calidad del hongo, sino también por su contexto ecológico y geográfico. La altitud y las especies de árboles sobre las que crece son factores que pueden influir en su perfil nutricional y propiedades organolépticas. La recolección de hongos silvestres como el “Qipatari” representa una práctica tradicional que puede contribuir a la sostenibilidad y a la economía local, al tiempo que ofrece un alimento nutritivo y potencialmente valioso para el mercado. Conocer sobre las condiciones de crecimiento y las carac-terísticas del hábitat es esencial para futuras inves-tigaciones sobre el cultivo y la conservación de este hongo, así como para su posible comercialización.
Resultados proporcionados por el Departamento de Filogenética y Evolución Molecular de la Universidad de Varsovia indicaron que se lograron obtener dos secuencias de alta calidad utilizando el cebador ITS1F (dos repeticiones de PCR). Las secuencias obtenidas correspondieron a una simili-tud de 88,30% - 89,97% con la especie Lentinus levis (Berk & M.A. Curtis) Murrill (comparación de BLASTn en NCBI para todas las bases de datos) y una similitud del 88,49% con Pleurotus cystidiosus var. formosensis (comparación BLASTn en NCBI, consultada sólo para materiales tipo).
De acuerdo con Rodríguez-Uña et al. (2019), un porcentaje de similitud inferior al 97% en la región ITS puede interpretarse como indicativo de que las muestras analizadas podrían corresponder a una especie no descrita previamente. En este caso, los resultados moleculares sugieren que el hongo “Qipatari” tiene una relación genética cercana con L. levis y P. cystidiosus var. formosensis, pero la baja similitud obtenida con estas especies conocidas podría ser evidencia de que el “Qipatari” podría constituir una nueva especie. Sin embargo, se requieren estudios adicionales, incluyendo análisis filogenéticos más detallados, así como la evalua-ción de características morfológicas y ecológicas, para corroborar esta hipótesis y determinar con precisión su clasificación taxonómica.
Es importante resaltar que el objetivo principal de este estudio no es profundizar en la taxonomía del hongo, sino documentar por primera vez su su cultivo exitoso en residuos lignocelulósicos y su potencial como hongo comestible. Este es el primer reporte que demuestra la domesticación de “Qipatari” y su posible uso alimenticio, no obstante, se deja abierta la posibilidad de futuras investi-gaciones que aborden con mayor profundidad su clasificación taxonómica y sus relaciones filogenéticas.
En cuanto al aislamiento micelial, únicamente dos placas Petri con medio de cultivo EMA tuvieron resultados positivos (Anexo 3) con un tiempo de reacción micelial de 72 horas, lo cual coincide con Gaitán-Hernández et al. (2006), quienes mencio-nan un tiempo de 2 a 3 días como un valor representativo para el inicio del crecimiento micelial en condiciones óptimas de cultivo. Ambas placas no tuvieron presencia de contaminantes visibles hasta el día 6, siendo nuevamente repicadas para observar que su desarrollo micelial fuera regular y constante. Tras este proceso, se seleccionó la cepa con mejor apariencia (pseudotejido algodonoso, tipo de borde regular, textura algodonosa y afelpada con densidad medianamente abundante), características que indican la alta calidad de una cepa para hongos comestibles, y por ser la primera en cubrir completamente la superficie.

Figura 2. Hongo silvestre “Qipatari”.
El análisis gráfico de la prueba de Tukey (Figura 3) destaca las diferencias significativas entre las formulaciones evaluadas. Se observa que F3 presenta las mayores diferencias medias en comparación con F5, F4 y F1, reforzando la hipótesis de que las propiedades químicas y estructurales del sustrato son determinantes en la colonización micelial del hongo “Qipatari”, como una mejor relación carbono-nitrógeno y una mayor degradabilidad de la lignina del aserrín de aliso en combinación con el rastrojo, lo que facilita el acceso a los nutrientes requeridos por el micelio (Kumla et al, 2020). La ausencia de diferencias significativas entre F1, F2 y F4, sugiere que, aunque el micelio del hongo puede invadir las mezclas con aserrín de eucalipto, éstas no le ofrecen las mismas condiciones óptimas que el aserrín de aliso, ya que éste conlleva una afinidad natural con el ambiente original donde el “Qipatari” fue hallado, lo cual mejora su adaptación al cultivo y facilita el proceso de domesticación. Estudios previos con otros basidiomicetos, como Pleurotus spp., también han reportado que la composición química de los sustratos lignocelulósicos influye significativa-mente en la eficiencia de colonización (Bermúdez-Savón et al., 2023; Delgado et al., 2024).
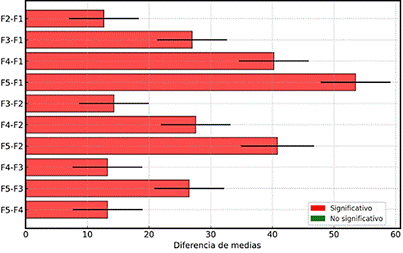
Figura 3. Prueba de Tukey: Comparaciones de diferencias de medias.
El bajo desarrollo del tratamiento F5 (Figura 4), determinó su exclusión de la etapa de fructificación para la obtención de carpóforos, dando a entender que el uso de chala de maíz como único compo-nente limita la capacidad de colonización micelial del “Qipatari”, a diferencia de otros hongos como P. ostreatus (Holgado et al., 2019). Esto podría estar relacionado con una baja biodisponibilidad de ciertos nutrientes clave para este hongo, lo cual coincide con investigaciones que destacan la importancia de utilizar mezclas de sustratos para optimizar el crecimiento de hongos comestibles y medicinales en general (Muswati et al., 2021).
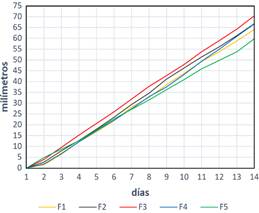
Figura 4. Crecimiento micelial lineal en las 5 formulaciones evaluadas. F1: 80% Aserrín de Aliso + 20% Rastrojo de trigo; F2: 80% Aserrín de eucalipto y 20% rastrojo de trigo; F3: 80% rastrojo + 20% aserrín de aliso; F4: 80% rastrojo y 20% de Aserrín de eucalipto; F5: Chala de maíz 100%.
Las formulaciones F2 y F4 presentaron problemas durante su desarrollo (Tabla 1), con interrupción de la incubación y sin fructificación, lo que sugiere que las formulaciones con aserrín de eucalipto no favorecieron el crecimiento y la producción de primordios; sin embargo, existen otros hongos como Lentinula edodes y Polyporus sanguinus (Omarini et al., 2009) que obtienen un buen desarrollo y fructificación en proporciones de aserrín de eucalipto hasta del 100%.
Tabla 1
Desarrollo del “Qipatari” en las diferentes formulaciones evaluadas durante el cultivo
|
Formu-lación |
Incubación |
Fructifi-cación |
Aparición de primordios (promedio) |
|
F1 |
Completada |
Sí |
41 días |
|
F2 |
Interrumpida |
No |
- |
|
F3 |
Completada |
Sí |
26 días |
|
F4 |
Completada |
No |
- |
Dónde: F1: 80% Aserrín de Aliso + 20% Rastrojo de trigo; F2: 80% Aserrín de eucalipto y 20% rastrojo de trigo; F3: 80% rastrojo + 20% aserrín de aliso; F4: 80% rastrojo y 20% de Aserrín de eucalipto.
Tras la única cosecha realizada en las formulacio-nes F1 y F3, al finalizar el experimento, ambas se mantuvieron en estado de fructificación con el objetivo de inducir una segunda cosecha. Este esfuerzo resultó en la obtención de una segunda oleada únicamente en tres bolsas de F1, que se produjo de manera gradual a lo largo de un mes. La ausencia de una segunda cosecha en F3 podría atribuirse al agotamiento de nutrientes, particularmente carbono, lo cual se vio exacerbado por el tamaño pequeño de las bolsas de prueba y por el menor porcentaje de aserrín en la formulación, que limitó la disponibilidad de lignina como fuente de carbono, a diferencia de F1 donde el 80% correspondía a aserrín de aliso.
La EB observada en F1 y F3 superó el 50% (Tabla 2), lo que las posiciona como producciones acep-tables y económicamente viables, según lo seña-lado por Guarín & Ramírez (2004). Se estima que estos valores podrían incrementarse significativa-mente si se empleara una mayor cantidad de sustrato por bolsa durante el cultivo.
La aparición tardía de primordios en F1 (41 días) en relación a F3 (26 días), podría indicar que la formulación F1 requiere más tiempo para alcanzar la madurez, probablemente por contener mayor porcentaje de aserrín que F3, lo que podría ser un factor limitante en términos de eficiencia de producción. Así mismo, el mayor porcentaje de rastrojo presente en F3, sugiere que su combi-nación con aserrín de aliso favorece un desarrollo más rápido, lo que podría ser un indicador de una mejor adaptación a estas condiciones.
Como se puede apreciar en la Figura 5, F1 muestra una predominancia de carpóforos en el tamaño G3 (10-14,9 cm) con un 75% de los carpóforos en este rango, mientras que F3 presenta una distribución más diversa, con carpóforos en los tamaños G2 (43,75%) y G3 (43,75%), y un pequeño porcentaje de carpóforos en G5 (6,25%). Esto podría sugerir que la formulación F3 produce una mayor variabilidad en el tamaño de los carpóforos, lo que podría ser beneficioso en términos de producción, ya que no todo el cultivo se concentra en un solo rango de tamaño. La formulación F3 muestra un peso fresco promedio de 134,59 g, significativa-mente mayor que el de F1 con 100,10 g (Tabla 2).
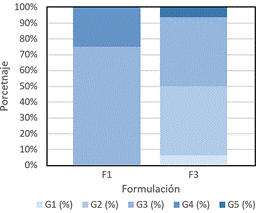
Figura 5. Porcentaje promedio de fructificaciones por formulación agrupadas por tamaño de pileo (G*). Dónde: G1< 4,9 cm, G2 = 5 -9,9 cm, G3 = 10 – 14, cm, G4 = 15 – 19,9 cm, G5 >20 cm. *Clasificación basada en Mata & Guzmán (1993).
Este aumento en el peso fresco puede explicarse por una mayor disponibilidad de nutrientes en F3, ya que la combinación de 80% rastrojo + 20% aserrín de aliso genera un sustrato más adecuado para el crecimiento micelial, lo que coincide con estudios previos sobre la influencia del sustrato en la biomasa de hongos (Sánchez & Montoya, 2020). Del mismo modo, F3 también supera a F1 en términos de Eficiencia Biológica, alcanzando 75,05% frente al 56% de F1, esto sugiere que la formulación F3 posibilita una mejor conversión del sustrato en biomasa, un aspecto crucial en la producción de hongos comestibles, como ha sido reportado en investigaciones sobre la optimización de formulaciones para mejorar la conversión biológica (Ahmed et al., 2024; Omarini et al., 2009; Albertó, 2008). Así mismo, Wan Mahari et al. (2020) mencionan que la paja de trigo, rica en celulosa, hemicelulosa y lignina, puede propor-cionar nutrientes para el desarrollo de la fructificación, obteniendo una mayor eficiencia biológica, mientras que el uso de aserrín como sustrato puede resultar en un rendimiento y desempeño más reducido, probablemente por el contenido proteico muy bajo del aserrín que es insuficiente para sustentar el crecimiento de algunas especies de hongos.
La tasa de productividad de F3 (0,8) también es más alta que la de F1 (0,52), lo que implica que F3 genera más biomasa por unidad de tiempo, haciendo más eficiente el uso del sustrato y potencialmente reduciendo los costos operativos. Este comportamiento es consistente con investigaciones que destacan la importancia de la formulación de sustratos para maximizar la productividad en el cultivo de hongos (Muswati et al., 2021), pues la eficiencia de la producción de hongos está estrechamente relacionada con la composición de nutrientes del sustrato de cultivo y el tipo de suplementos añadidos (Kumar et al., 2022). El uso de rastrojo como sustrato ha mostrado tener un impacto positivo en la aceleración del proceso de fructificación para hongos comestibles (Archana et al., 2024). En general los residuos leñosos (aserrín) y agrícolas (rastrojo) difieren en características químicas y estructurales, por lo que la biomasa agrícola típicamente tiene contenidos más bajos de lignina y carbohidratos que la biomasa leñosa, además de diferir estructuralmente entre sí (Yin et al., 2025); de acuerdo a ello se infiere que la formulación F3 siendo la mejor y la más óptima, tuvo el menor Ciclo de cultivo posiblemente por la alta cantidad de hemicelulosa y celulosa, presente en el compo-nente mayoritario que fue el rastrojo de trigo (80%), lo cual indica una degradación más rápida por parte del hongo; a diferencia de la fórmula F1 que tenía mayor proporción de aserrín de aliso que implica un mayor contenido de lignina y por ende mayor tiempo de degradación (Chang & Miles, 1989). El rendimiento de F3 (25,91%), superior al de F1 (19,6%), indica que F3 utiliza de manera más efectiva el sustrato para producir biomasa útil. Este tipo de datos es relevante para la industria de los hongos, donde optimizar el rendimiento es clave para mejorar la rentabilidad de la producción ya que las diferentes especies de hongos cultivados requieren de sustratos con características especí-ficas, tanto en la formulación de los ingredientes como en el proceso de elaboración (Zied et al., 2024).
El ciclo de cultivo de F3 es 92 días, más corto que el de F1 (107 días). Esta reducción en el ciclo puede ser una ventaja competitiva, al aumentar considerablemente la productividad permitiendo una mayor rentabilidad (Jaramillo & Albertó, 2019). Los carpóforos cultivados en las formulaciones F1 y F3 (Tabla 3), muestran un incremento proteico significativo de 24,92% y 24,7% respectivamente, en comparación con el hongo silvestre (15.9%) lo cual puede atribuirse a las condiciones de cultivo que optimizan la disponibilidad de nutrientes en los sustratos (Roncero et al., 2015).
Tabla 2
Parámetros de productividad del hongo comestible “Qipatari” por formulación
|
Formulación |
Promedio de peso fresco (g) |
Eficiencia Biológica (%) |
Tasa de Productividad |
Rendimiento (%) |
Ciclo de cultivo (días) |
|
F1 |
100,10 |
56 |
0,52 |
19,6 |
107 |
|
F3 |
134,59 |
75,05 |
0,8 |
25,91 |
92 |
Tabla 3
Comparación del contenido nutricional de los carpóforos silvestres y cultivados del hongo comestible “Qipatari” con otros hongos comerciales
|
Componentes nutricionales |
“Qipatari” silvestre |
“Qipatari” cultivado (F1) |
“Qipatari” cultivado (F3) |
*Lentinula edodes (Shiitake) |
*Pleurotus ostreatus (hongo ostra) |
|
Humedad % |
9,9 |
10,3 |
12,62 |
- |
- |
|
Proteína % |
15,9 |
24,92 |
24,7 |
13,4-17,5 |
10,5-30,4 |
|
Grasa % |
2,13 |
1,91 |
1,88 |
4,9-8 |
1,6-2,2 |
|
Ceniza % |
8,35 |
7,78 |
7,79 |
3,3-7,0 |
6,1-9,8 |
|
Fibra % |
13,8 |
12 |
11,3 |
7,3-8,0 |
7,5-8,7 |
|
Carbohidratos % |
68,32 |
55,09 |
53,01 |
67,5-81,8 |
57,6-81,8 |
|
Ca mg/100 |
72,7 |
65,7 |
64,2 |
50 |
10 |
|
Mg mg/100 |
110,2 |
88 |
86,4 |
1550 |
2000 |
|
Fe mg/100 |
10,7 |
11,42 |
11,6 |
33 |
54 |
|
P mg/100 |
697,4 |
616,8 |
622,3 |
8700 |
13900 |
|
Zn mg/100 |
11,9 |
10,8 |
10 |
92 |
83 |
|
K mg/100 |
1240,4 |
1040 |
996,2 |
26700 |
3730 |
Laboratorio de Análisis Químico – UNSAAC, *Chang & Miles (2004).
Comparado con P. ostreatus (10,5% – 30,4%) y L. edodes (13,4% – 17,5%), los niveles de proteína del Qipatari son competitivos, consolidándolo como una excelente fuente proteica para dietas vegetarianas o como alternativa a los productos animales; más aun considerando que las proteínas de los hongos han atraído más atención en los últimos tiempos, debido a los menores costos de producción y la reducción de los problemas ambientales en comparación con las proteínas animales y vegetales, a la vez que poseen algunas cualidades únicas que las convierten en fuentes de proteínas alternativas adecuadas en el futuro (Ayimbila & Keawsompong, 2023). Así mismo, el contenido proteico de los carpóforos silvestres posiciona al “Qipatari” como una fuente proteica prometedora, similar a hallazgos en hongos nativos de la Patagonia que destacan altos contenidos proteicos en especies como Lepista nuda (30,3%) (Rugolo et al., 2022).
El contenido graso del “Qipatari” se sitúa entre 1,88% – 2,13%, semejante a lo reportado para P. ostreatus (1,6% – 2,2%) y significativamente menor que el de L. edodes (4,9% – 8%). Este perfil lipídico, junto con la presencia de ácidos grasos insaturados como el linoleico (3,288 mg/100 g), refuerza su rol como alimento saludable, con beneficios potenciales en la regulación del colesterol y la salud cardiovascular (Kalac, 2013).
Así mismo, el contenido de fibra en el “Qipatari” (11,3% – 13,8%) superan a los de P. ostreatus y L. edodes (7,3% – 8,7%), posicionándolo como un alimento funcional importante para mejorar la salud digestiva y prevenir enfermedades metabóli-cas (Albertó, 2008; Rugolo et al., 2022). La reduc-ción de la presencia de carbohidratos en los hongos cultivados podría deberse al metabolismo durante el proceso de domesticación, esto es similar a lo observado en otros hongos nativos, donde las cepas recién domesticadas, y las cultivadas comercial-mente, mostraban perfiles nutricionales similares, con alteraciones metabólicas durante las distintas etapas del desarrollo. Esto sugiere que el proceso de cultivo podría estabilizar ciertos rasgos metabólicos en los hongos, como los carbohidratos (Fu et al., 2022; Hashemian et al., 2019).
Del mismo modo, el hongo presenta un contenido notable de minerales esenciales (Tabla 3), destacando niveles significativos de potasio (996,2 – 1240,4 mg/100 g), fósforo (616,8 – 697,4 mg/100 g) y hierro (11,42 – 11,6 mg/100 g). Estos valores son superiores a los de otros hongos como P. ostreatus. Así mismo, el contenido de potasio del “Qipatari” lo hace relevante para el manejo de la presión arterial, mientras que sus altos niveles de hierro pueden contribuir a combatir la anemia (Teja et al., 2023).
Por otro lado, el perfil de aminoácidos del “Qipatari” (Tabla 4) incluye 8 de los 10 aminoácidos esencia-les, destacando altos niveles de alanina, leucina y lisina. La presencia significativa de arginina y ácido glutámico refuerza sus propiedades funcionales, mientras que los azúcares libres como fructosa y glucosa contribuyen al valor energético del hongo.
Tabla 4
Aminoácidos esenciales, no esenciales, azúcares y ácidos grasos presentes en los carpóforos silvestres del hongo comestible “Qipatari”
|
|
Tipo |
Aminoácido mg/100g |
|
Aminoácidos Esenciales
|
Histidina |
53,7 |
|
Treonina |
70,80 |
|
|
Alanina |
268,1 |
|
|
Valina |
136,2 |
|
|
Metionina |
0,0 |
|
|
Fenilalanina |
60,1 |
|
|
Isoleucina |
69,5 |
|
|
Leucina |
123,0 |
|
|
Lisina |
118,4 |
|
|
Triptófano |
nd |
|
|
Aminoácidos No Esenciales
|
Ácido aspártico |
88,1 |
|
Acido |
274,0 |
|
|
Glutámico |
||
|
Serina |
324,9 |
|
|
Glicina |
293,2 |
|
|
Arginina |
569,1 |
|
|
Tirosina |
210,0 |
|
|
Cistina |
0,0 |
|
|
Prolina |
0,0 |
|
|
Carbohidratos g/100g |
Fructosa |
11,100 |
|
Glucosa |
26,949 |
|
|
Ácidos grasos mg/100 g |
Ácido palmítico |
0,942 |
|
Ácido oleico |
3,218 |
|
|
Ácido Linoleico |
3,288 |
La predominancia de ácidos grasos polinsaturados como el oleico y linoleico subraya su papel como alimento funcional ayudando a reducir los niveles de colesterol LDL (colesterol “malo”) y promovien-do un perfil lipídico saludable (Sande et al., 2019). Todos estos nutrientes hacen del hongo una opción atractiva para quienes buscan mejorar su ingesta nutricional a través de fuentes vegetales.
CONCLUSIONES
La recolección de carpóforos del hongo comestible “Qipatari” en la localidad de Río Blanco, Apurímac, permitió identificar su preferencia por sustratos lignocelulósicos, específicamente troncos muertos de A. acuminata y M. oreophila. El análisis molecular preliminar reveló similitudes del 88,30-89,97% con Lentinus levis y del 88,49% con P. cystidiosus var. formosensis, lo que sugiere una relación genética cercana con estas especies. Sin embargo, estos porcentajes de similitud, inferiores al 97%, podrían indicar que se trata de una especie no descrita previamente, como lo sugieren los estándares de delimitación taxonómica menciona-dos en la literatura.
El exitoso aislamiento micelial y su cultivo hasta la obtención de cuerpos fructíferos, utilizando mezclas de sustratos lignocelulósicos, demuestran que el “Qipatari” es un hongo susceptible a la domesticación y producción comercial. La formulación F3, compuesta por 80% de rastrojo de trigo y 20% de aserrín de aliso, fue la más efectiva, con una alta eficiencia biológica y una óptima conversión del sustrato en biomasa, sugiriendo que su combinación de nutrientes optimiza el rendimiento del cultivo.
Además, los carpóforos silvestres y los obtenidos en cultivo, presentaron un perfil nutricional prometedor, con altos niveles de proteínas, minerales esenciales como potasio y hierro, y una notable cantidad de fibra, superando a otros hongos comestibles como P. ostreatus y L. edodes. Estos resultados destacan al “Qipatari” como una fuente proteica funcional con potencial para dietas vegetarianas y beneficios para la salud cardiovascular y digestiva, posicionándolo como una alternativa alimenticia saludable y competitiva en el mercado de hongos comestibles.
Finalmente, aunque este trabajo constituye el primer reporte de comestibilidad y cultivo de este hongo nativo, es imprescindible realizar estudios complementarios, incluyendo un análisis mole-cular filogenético más detallado, su descripción taxonómica y la evaluación de sus propiedades bioquímicas y biotecnológicas, para comprender plenamente su potencial y relevancia científica.
AGRADECIMIENTOS
Los autores expresan su agradecimiento a Don Edgar Nicolás Laura, y a sus sobrinos Hans y Yaneth por su apoyo como guías de campo y por sus conocimientos acerca de la colecta y consumo del “Qipatari”, así mismo a las autoridades y pobladores de las localidades de Río Blanco y Sapichaca, a Yeshica Huamán Pérez y Marleni Huamán Pérez por la hospitalidad brindada. Nuestro reconocimiento a la Dra. Marta Wrzosek (Departament of Molecular Phylogenetics and Evolution, University of Warsaw) por su invaluable apoyo en el análisis molecular. Al Programa Yachaininchis Wiñarinanpaq por el financiamiento de Tesis de Posgrado del Convenio UNSAAC-FONDECYT de la Universidad Nacional de San Antonio Abad del Cusco, por financiar parte de esta investigación.
REFERENCIAS BIBLIOGRÁFICAS
Aguilar-Pumahuillca, F., Huamán-Huamán, H., & Holgado-Rojas, M. (2019). Caracterización de Pleurotus sp. aislado de la Comunidad Nativa de Korimani, Centro Poblado de Kiteni-Echarate, La Convención, Cusco, Perú. Ecología Aplicada, 18(1), 45-50. https://doi.org/10.21704/rea.v18i1.1305
Ahmed, R., Niloy, M. A. H. M., Islam, M. S., Reza, M. S., Yesmin, S., Rasul, S. B., & Khandakar, J. (2024). Optimizing tea waste as a sustainable substrate for oyster mushroom (Pleurotus ostreatus) cultivation: A comprehensive study on biological efficiency and nutritional aspect. Frontiers in Sustainable Food Systems, 7. https://doi.org/10.3389/fsufs.2023.1308053
Albertó, E. (2017). Naturally occurring strains of edible mushrooms: A source to improve the mushroom industry. En C. Z. Diego & A. Pardo-Giménez (Eds.), Edible and medicinal mushrooms (cap. 19). https://doi.org/10.1002/9781119149446.ch19
Albertó, E. (2008). Cultivo intensivo de los hongos comestibles: Cómo cultivar champiñones, gírgolas, shiitake y otras especies (1ª ed.). Editorial Hemisferio Sur.
Alim, Y., Sidhoum, W., & Dib, S. (2023). First record of the edible mushroom Lepista sordida in Western Algerian Forest: Nutritional value and physicochemical parameters of mycelial culture. Journal of Fungi, 9(8), 858. https://doi.org/10.3390/jof9080858
Arana-Gabriel, Y., Burrola-Aguilar, C., Alcalá-Adán, A., Zepeda, C., & Estrada-Zúñiga, M. (2020). Mycelial growth of the edible wild mushrooms Floccularia luteovirens in different culture mediums and pH. AgroProductividad, 13, 33-38. https://doi.org/10.32854/agrop.v13i10.1745
Ayimbila, F., & Keawsompong, S. (2023). Nutritional quality and biological application of mushroom protein as a novel protein alternative. Current Nutrition Reports, 12(4), 281-298. https://doi.org/10.1007/s13668-023-00468-x
Cano, A., & Romero, L. (2016). Valor económico, nutricional y medicinal de hongos comestibles silvestres. Revista Chilena de Nutrición, 43(1), 75-80.
Chang, S. T., & Miles, P. G. (2004). Mushrooms: Cultivation, nutritional value, medicinal effect, and environmental impact. CRC Press.
Chang, S. T., & Miles, P. G. (1989). Edible mushrooms and their cultivation. CRC Press.
Chimey, C., & Holgado, M. E. (2010). Los hongos comestibles silvestres y cultivados en Perú. En D. Martínez-Carrera, N. Curvetto, M. Sobal, P. Morales, & V. M. Mora (Eds.), Hacia un desarrollo sostenible del sistema de producción-consumo de los hongos comestibles y medicinales en Latinoamérica: Avances y perspectivas en el siglo XXI (pp. 381-395). Red Latinoamericana de Hongos Comestibles y Medicinales-COLPOS-UNS-CONACYT-AMC-UAEM-UPAEP-MINAP.
Curvetto, N., Figlas, D., & Delmastro, S. (2002). Sunflower seed hulls as substrate for the cultivation of shiitake mushrooms. HortTechnology, 12(4), 652-655. https://doi.org/10.21273/HORTTECH.12.4.652
Dávila-Arenas, C., Sulca-Quispe, L., & Pavlich-Herrera, M. (2018). Estudio Etnomicológico de la micobiota comestible en dos Comunidades Nativas de la Cuenca Alto Madre de Dios, Reserva Biósfera Del Manu. Sagasteguiana, 1(1), 121-130.
Delgado, N., Miranda, V., Barros, J., Isla, M., & Fracchia, S. (2024). Exploring local lignocellulosic substrates for the production of edible mushrooms in Northwestern Argentina. Lilloa, 61(2), 317-339. https://doi.org/10.30550/j.lil/1973
De Jesús-Rivera, L., Álvarez-Sánchez, M. E., Ramírez-Pérez, F., & Maldonado-Torres, R. (2022). Prickly pear glochids for commercial production of oyster mushroom. Revista Mexicana de Ciencias Agrícolas, 13(8), 1445–1454. https://doi.org/10.29312/remexca.v13i8.3351
Fernández, F. (2005). Manual práctico de producción comercial de champiñón. Guadalajara, Jalisco, México.
Fu, Y. P., Dai, Y. T., Chethana, K. W. T., Li, Z. H., Sun, L., Li, C. T., Yu, H. L., Yang, R. H., Tan, Q., Bao, D. P., Deng, Y. J., Wang, S. X., Wang, Y. F., Tian, F. H., Qi, L. L., Shu, L. L., Jia, P. S., Chen, L. C., Chen, M. Y., Hu, Q. X., Tan, H., Song, T. T., Zhang, Z. W., Bonito, G., Zervakis, G. I., Xiao, S. J., Hyde, K. D., Li, Y., & Yuan, X. H. (2022). Large-scale genome investigations reveal insights into domestication of cultivated mushrooms. Mycosphere, 13(2), 86–133. https://doi.org/10.5943/mycosphere/si/1f/4
Gaitán-Hernández, R., Salmones, D., Pérez, R., & Mata, G. (2006). Manual práctico del cultivo de setas: Aislamiento, siembra y producción. Xalapa: Instituto de Ecología.
García, M. (2007). Cultivo de setas y trufas. Mundi-Prensa Libros.
Holgado, M. E., Aranzabal, R., Lazarte, R., Quispe, A., Pérez, K. A., Aguilar, F. B., & Aguilar, F. (2019). Cultivo de Pleurotus spp. y Lentinula edodes bajo condiciones artesanales en comunidades campesinas de la Región Cusco. Revista de Ecología Aplicada, 18(2). https://doi.org/10.21704/rea.v18i2.1331
Jaramillo, S., & Albertó, E. (2019). Incremento de la productividad de Pleurotus ostreatus mediante el uso de inóculo como suplemento. Scientia Fungorum, 49, e1243. https://doi.org/10.33885/sf.2019.49.1243
Jasinska, A. (2023). Sustainability of mushroom cultivation systems. Horticulturae, 9(11), 1191. https://doi.org/10.3390/horticulturae9111191
Kalac, P. (2013). A review of chemical composition and nutritional value of wild-growing and cultivated mushrooms. Journal of the Science of Food and Agriculture, 93, 209-218. https://doi.org/10.1002/jsfa.5960
Kumar, P., Eid, E. M., Al-Huqail, A. A., Širić, I., Adelodun, B., Abou Fayssal, S., Valadez-Blanco, R., Goala, M., Ajibade, F. O., Choi, K. S., & Kumar, V. (2022). Kinetic studies on delignification and heavy metals uptake by shiitake (Lentinula edodes) mushroom cultivated on agro-industrial wastes. Horticulturae, 8(4), 316. https://doi.org/10.3390/horticulturae8040316
Kumawat, A., Wasnikar, A. R., Kumawat, H., Kumawat, G., Kumawat, K. C., & Rani, K. (2024). Insight into the effect of crop straw on production of oyster mushroom (Pleurotus ostreatus). Biological Forum – An International Journal, 16(7), 300-303.
Kumla, J., Suwannarach, N., Sujarit, K., Penkhrue, W., Kakumyan, P., Jatuwong, K., Vadthanarat, S., & Lumyong, S. (2020). Cultivation of mushrooms and their lignocellulolytic enzyme production through the utilization of agro-industrial waste. Molecules, 25, 2811. https://doi.org/10.3390/molecules25122811
Lallawmsanga, A., & Carrasco, B. (2022). Diversity of the fungi kingdom: Molecular tools to distinguish mushrooms considered safe and unsafe for human health. En D. Stojković & L. Barros (Eds.), Edible fungi: Chemical composition, nutrition, and health effects (Chapter 1, pp. 1-22). Food Chemistry, Function and Analysis (Vol. 36). The Royal Society of Chemistry.
López, A. (2007). Manual de producción de micelio de hongos comestibles. Instituto de Genética Forestal-Universidad de Veracruz, México.
Mata, G., & Guzmán, G. (1993). Cultivation of Lentinus boryanus in wood shavings in Mexico. Cryptogamic Botany, 4, 47–49.
Muswati, C., Simango, K., Tapfumaneyi, L., Mutetwa, M., & Ngezimana, W. (2021). The effects of different substrate combinations on growth and yield of oyster mushroom (Pleurotus ostreatus). International Journal of Agronomy, 2021, Art. 9962285. https://doi.org/10.1155/2021/9962285
Mushworld. (2005). Handbook of cultivators - Shiitake. Seúl, Corea del Sur: Editorial Haeng-oon.
Naim, L., Alsanad, M., El Sebaaly, Z., Shaban, N., Abou Fayssal, S., & Sassine, Y. (2020). Variation of Pleurotus ostreatus (Jacq. ex Fr.) P. Kumm. (1871) performance subjected to different doses and timings of nano-urea. Saudi Journal of Biological Sciences, 27, 2794–2801. https://doi.org/10.1016/j.sjbs.2020.03.019
Omarini, A., Lechner, B. E., & Albertó, E. (2009). Polyporus tenuiculus: A new naturally occurring mushroom that can be industrially cultivated on agricultural waste. Journal of Industrial Microbiology & Biotechnology, 36, 635–642. https://doi.org/10.1007/s10295-009-0530-2
Okuda, Y. (2022). Sustainability perspectives for future continuity of mushroom production: The bright and dark sides. Frontiers in Sustainable Food Systems, 6, 1026508. https://doi.org/10.3389/fsufs.2022.1026508
Pavlich, M. R. (2001). Los hongos comestibles del Perú. Revista de Ciencias Biológicas BIOTA, 18, 3-19.
Pérez-Moreno, J., Mortimer, P. E., Xu, J., Karunarathna, S. D. O., & Li, H. (2021). Global perspectives on the ecological, cultural and socioeconomic relevance of wild edible fungi. Studies in Fungi, 6(1), 408-424. https://doi.org/10.5943/sif/6/1/31
Ramírez-Terrazo, A., Montoya, A., & Kong, A. (2021). Conocimiento micológico tradicional en dos comunidades aledañas al Parque Nacional Lagunas de Montebello, Chiapas, México. Scientia Fungorum, 51, e1321. https://doi.org/10.33885/sf.2021.51.1321
Raza, M. H., Abid, M., Faisal, M., Yan, T., Akhtar, S., & Adnan, K. M. M. (2022). Environmental and health impacts of crop residue burning: Scope of sustainable crop residue management practices. International Journal of Environmental Research and Public Health, 19(8), 4753. https://doi.org/10.3390/ijerph19084753
Roncero, I. (2015). Propiedades nutricionales y saludables de los hongos. Centro Tecnológico de Investigación del Champiñón de La Rioja (CTICH).
Rugolo, M., Mascoloti Spréa, R., Dias, M. I., Pires, T. C. S. P., Añibarro-Ortega, M., Barroetaveña, C., Caleja, C., & Barros, L. (2022). Nutritional composition and bioactive properties of wild edible mushrooms from native Nothofagus Patagonian forests. Food, 11(21), 3516. https://doi.org/10.3390/foods11213516
Salmones, D., Gaitán-Hernández, R., Pérez, R., & Guzmán, G. (1997). Studies on genus Pleurotus. VIII. Interaction between mycelial growth and yield. Revista Iberoamericana de Micología, 14, 173-176.
Sánchez, Ó. J., & Montoya, S. (2020). Assessment of polysaccharide and biomass production from three white-rot fungi by solid-state fermentation using wood and agro-industrial residues: A kinetic approach. Forests, 11(10), 1055. https://doi.org/10.3390/f11101055
Sánchez, J. E., & Royse, D. (2001). La biología y el cultivo de Pleurotus spp. El Colegio de la Frontera Sur, Carretera Panamericana y Periférico Sur s/n, Barrio de María Auxiliadora
Sande, D., Pereira de Oliveira, G., Fidelis, M. A., Martins, B. A., Lima, M. T. N. S., & Takahashi, J. A. (2019). Edible mushrooms as a ubiquitous source of essential fatty acids. Food Research International, 125, 108524. https://doi.org/10.1016/j.foodres.2019.108524
Stamets, P., & Chilton, J. (1983). The mushroom cultivator: A practical guide to growing mushrooms at home. Agarikon Press.
Teja, K. S., Suruchi, Rai, U., Kumar, M., Mohanty, O., Roy, J., & Meshram, S. (2023). Mushrooms: A potential option in the management of deficiency and diseases in humans. Journal of Pure and Applied Microbiology, 17(2), 749-760. https://doi.org/10.22207/JPAM.17.2.55
Tekeste, N., Dessie, K., Taddesse, K., & Ebrahim, A. (2020). Evaluation of different substrates for the yield and yield attributes of oyster mushroom (Pleurotus ostreatus) in the crop-livestock system of northern Ethiopia. The Open Agriculture Journal, 14(1), 30–40. https://doi.org/10.2174/1874331502014010030
Trutmann, P., Luque Luque, A., Holgado Rojas, M., Quispe Pelaez, A., Callañaupa Auccapuma, M., López Mesones, M., & Pavlich Herrera, M. (2022). The mushrooms of Andean Peru: Part 1. Order Agaricales: Families Agaricaceae to Lyophyllaceae (1st ed.). Global Mountain Action Publications, Orselina, Switzerland. ISBN-13: 979-8987167014.
Wan Mahari, W. A., Peng, W., Nam, W. L., Yang, H., Lee, X. Y., Lee, Y. K., Liew, R. K., Ma, N. L., Mohammad, A., Sonne, C., Van Le, Q., Show, P. L., Chen, W.-H., & Lam, S. S. (2020). A review on the valorization of oyster mushrooms and the waste generated in the mushroom farming industry. Journal of Hazardous Materials, 400, 123156. https://doi.org/10.1016/j.jhazma.2020.12
Yin, Y., Chen, B., Xu, S., Zuo, J., Xu, Y., Xiong, S., & Chen, F. (2025). Investigation of crop straw for edible and medicinal fungi cultivation: Assessment of lignocellulose preprocessing and spent substrate biofuel properties. Industrial Crops and Products, 223, 120004. https://doi.org/10.1016/j.indcrop.2024.120004
Zariquiey, R., & Córdova, G. (2008). Qayna, Kunan, Paqarin: Una introducción práctica al quechua chanca. Estudios Generales Letras - Pontificia Universidad Católica del Perú.
Zied, D. C., Pardo, J. E., Noble, R., & Pardo-Giménez, A. (2024). Efficiency of mushrooms for food production—Fundamental strategic decision-making. Journal of Food Composition and Analysis, 125, 105734. https://doi.org/10.1016/j.jfca.2024.105734
ANEXOS
Anexo 1
Tratamientos experimentales
|
Tratamiento |
Fórmula de Sustrato |
% Aserrín |
% Rastrojo |
% Chala de Maíz |
% Yeso |
Humedad (%) |
|
F1 |
Aserrín de Aliso + Rastrojo de Trigo |
80% |
20% |
0% |
1% |
65% |
|
F2 |
Aserrín de Eucalipto + Rastrojo de Trigo |
80% |
20% |
0% |
1% |
65% |
|
F3 |
Rastrojo de Trigo + Aserrín de Aliso |
20% |
80% |
0% |
1% |
65% |
|
F4 |
Rastrojo de Trigo + Aserrín de Eucalipto |
20% |
80% |
0% |
1% |
65% |
|
F5 |
Chala de Maíz |
0% |
0% |
100% |
1% |
65% |
Anexo 2
Ubicación geográfica del lugar de colecta
Anexo 3
Desarrollo del hongo Qipatari durante el aislamiento
|
Medio |
Placa |
Día 2 |
Día 3 |
Día 4 |
Día 5 |
Día 6 |
|
PDA |
1 |
0 |
0 |
- |
- |
- |
|
2 |
bacterias |
++ |
++ |
se detuvo |
- |
|
|
3 |
0 |
++ |
++ |
++ |
++ |
|
|
EMA |
4 |
0 |
0 |
0 |
- |
- |
|
5 |
0 |
++ |
+++ |
++ |
+++ |
|
|
6 |
0 |
bacterias |
+ |
- |
- |
Dónde 0: Desarrollo negativo; +: Desarrollo Regular; ++: Desarrollo bueno; +++: Desarrollo muy bueno; -: Se descartó.