 |
INTRODUCTION
Harnessing Chara globularis: Sustainable biomass harvest for water quality management and organic fertilizer production in Lake Titicaca
Aprovechamiento de Chara globularis: Cosecha sostenible de biomasa para el manejo de la calidad del agua y la producción de fertilizante orgánico en el lago Titicaca
Angel Canales-Gutiérrez1, 2 *; Marisol Chambi-Alarcón1, 2; Gelvi Canales-Manchuria3
1 Universidad Nacional del Altiplano de Puno. Facultad de Ciencias Biológicas. Programa de Ecología, Av. Floral 1153, Puno, Perú.
2 Instituto de Investigaciones Alto Altoandinas, Av. Floral 1153, Puno, Perú.
3 Universidad Nacional Jorge Basadre Grohmann. Escuela Profesional de Ingeniería Ambiental, Miraflores S/N, Tacna, Perú.
* Corresponding author: acanales@unap.edu.pe (A. Canales).
ORCID de los autores:
A. Canales-Gutiérrez: https://orcid.org/0000-0002-3096-1705 M. Chambi-Alarcón: https://orcid.org/0000-0001-9639-6385
G. Canales-Manchuria: https://orcid.org/0000-0001-8099-4276
ABSTRACT
Chara globularis, a submerged macroalga dominant in high-Andean lakes, generates controversial effects on freshwater ecosystems. Although its dense coverage alters water chemistry and reduces biodiversity, its abundant biomass offers an opportunity for sustainable use. This study assessed its seasonal dynamics, ecological impact, and productive potential in Lake Titicaca (Puno, Peru), from February to July 2024. In zones with and without Chara, a maximum biomass of 705,600 kg/ha and an average of 24,800 kg/ha across 43,600 ha were recorded, totaling over 1.08 million tons. Strong positive correlations were found with pH (r = 0.91) and electrical conductivity, along with a reduction in dissolved oxygen (3.7 mg/L), indicating potential hypoxia. The richness of macroinvertebrates and macroalgae was lower in Chara-dominated areas, reflecting ecological homogenization. However, its nutrient profile revealed high levels of potassium, phosphorus, nitrogen, and organic matter, supporting its use in compost production. A sustainable harvest of 60% (648,768 t) is proposed, as a strategy that combines ecological conservation with circular bioeconomy. Thus, C. globularis emerges as both a management challenge and a renewable resource for organic agriculture and environmental restoration.
Keywords: biomass; sustainable harvest; macrophyte; water quality; circular economy.
RESUMEN
Chara globularis, una macroalga sumergida dominante en lagos altoandinos genera efectos controversiales en los ecosistemas dulceacuícolas. Aunque su densa cobertura modifica la química del agua y reduce la biodiversidad, su abundante biomasa representa una oportunidad para el aprovechamiento sostenible. Este estudio evaluó su dinámica estacional, impacto ecológico y potencial productivo en el lago Titicaca (Puno, Perú), entre febrero y julio de 2024. En zonas con y sin Chara, se halló una biomasa máxima de 705600 kg/ha y un promedio de 24800 kg/ha en 43600 ha, equivalente a más de 1,08 millones de toneladas. Se registraron correlaciones positivas con pH (r = 0,91) y conductividad eléctrica, y una reducción del oxígeno disuelto (3,7 mg/L), indicando posible hipoxia. La riqueza de macroinvertebrados y macroalgas fue menor en zonas dominadas por Chara, evidenciando homogeneización ecológica. Sin embargo, su perfil nutricional mostró altos niveles de potasio, fósforo, nitrógeno y materia orgánica, lo que respalda su uso en compost. Se propone una cosecha sostenible del 60% (648768 t), como estrategia que combina conservación ecológica y bioeconomía circular. Así, C. globularis se presenta como un reto de manejo, pero también como un recurso renovable para la agricultura orgánica y la restauración ambiental.
Palabras clave: biomasa; cosecha sostenible; macrófita; calidad del agua; economía circular.
Recibido: 03-03-2025.
Aceptado: 09-06-2025.
|
Collot et al. (1983) indicate that Characeae occupy an area of 436 km² in the smaller basin of the lake, representing more than 60% of the vegetation-covered area. Along a transect between Cojata Island and Taraco Point, they extend for over 40 km on a bed covered with Chara, confirming the abundance of this plant group in the lake
The rapid expansion of Chara globularis in high-altitude aquatic ecosystems, such as Lake Titicaca, has prompted ecological concern due to its effects on water chemistry and biodiversity. Dense mats of Chara are associated with decreased dissolved oxygen and increased conductivity, resulting from restricted gas exchange and metabolic byproduct accumulation (Hutorowicz & Dziedzic, 2008; Gette et al., 2014). Its dominance is further facilitated by its affinity for alkaline environments, as indicated by positive correlations between biomass and pH (Rodrigo et al., 2015; Pełechaty et al., 2015).
The displacement of native aquatic flora through competition for light and space, coupled with potential allelopathic interactions, diminishes ecosystem resilience and homogenizes habitats (Bakker et al., 2010; Hutorowicz & Dziedzic, 2008). Although tolerant species like Hyalella sp. may persist, more sensitive taxa often decline, affecting the ecological integrity of these systems.
Despite these negative impacts, C. globularis biomass offers potential value. Its capacity to accumulate nitrogen, phosphorus, and potassium suggests possible applications in organic agriculture (Asaeda et al., 2014; Borges et al., 2023). Therefore, a management strategy based on sustainable biomass harvesting could both mitigate ecological damage and promote resource reuse. This dual-function approach aligns with circular bioeconomy principles and supports conservation efforts.
Diversos estudios han demostrado el papel ecológico y productivo de Chara globularis y otras macrófitas en ambientes acuáticos. Thakur et al. (2022) señalaron su potencial como biomasa para digestión anaerobia, alcanzando producciones de hasta 605 L CH₄/kg SV en co-digestión con lodos. En un lago de los Países Bajos, Van-Zuidam et al. (2022) observaron que Chara alcanzó una cobertura del 90–95% y resistió el pastoreo de cisnes, manteniendo estructura funcional. En cuanto a reproducción, Van-Onsem et al. (2018) evidenciaron que la producción de oósporas de Chara no se vio afectada por cobertura de Lemna minor, a diferencia de otras especies como Potamogeton pusillus. Maredová et al. (2021) evaluaron efectos alelopáticos y hallaron que solo 15 de 532 ensayos inhibieron cianobacterias, entre ellos Chara globularis, indicando un efecto limitado. Finalmente, Brzozowski & Pełechaty (2024) reportaron que la dominancia otoñal de Lychnothamnus barbatus frente a la competencia estival de Myriophyllum spicatum preserva biodiversidad en lagos templados, mostrando respuestas fenológicas adaptativas al cambio climático.
Several studies have revealed that species of the genus Chara exhibit remarkable ecological, morphological, and physiological adaptations to diverse aquatic environments. Laffont-Schwob et al. (2015) evaluated the capacity of Chara spp. to extract heavy metals in Mediterranean highway retention ponds, concluding that although their biomass is lower than that of Typha spp., they show comparable efficiencies in accumulating metals such as Cd, Cu, Zn, and Pb—particularly Chara vulgaris. Sviben et al. (2018) highlighted significant heterogeneity in photosyn-thetic pigments, calcite encrustation, and epiphyton communities in Chara subspinosa and C. globularis from the marl lake Prošće. Notably, C. globularis produced substantial biomass in deeper zones, suggesting a strong potential for sustainable harvesting. This abundant biomass, enriched with organic content and calcium carbonate deposits, holds promising value as a raw material for compost-based organic fertilizer, contributing to circular bioeconomy models in freshwater systems. Rybak et al. (2017) further demonstrated the sensitivity of Chara hispida to iron-based coagulants used in lake restoration, which can inhibit growth and reproductive structures by reducing light penetration and altering pH. These findings reinforce the importance of protecting viable Chara habitats for both ecological balance and resource utilization.
From a biogeographical and paleoecological perspective, Chemeris et al. (2020) documented eight species of charophytes in extreme permafrost environments of northeastern Asia, such as Chara globularis and C. contraria, which exhibit flexible life cycles enabling them to persist in harsh, short-season environments. Likewise, et al. (2023) identified eleven charophyte species from Early Pleistocene deposits in Turkey, with C. vulgaris, C. globularis, and C. hispida as dominant taxa, revealing transitions from palustrine to oligotrophic lake environments driven by glacial climate fluctuations. In another paleo-botanical study, Demirci et al. (2025) analyzed Late Miocene charophyte fossils from Central Anatolia, documenting species such as Lychnothamnus duplicicarinatus and Nitellopsis etrusca, whose stratigraphic presence reflects historical lake-level dynamics and regional tectonic shifts.
Finally, Gruszka et al (2023) investigated the controversial status of Chara connivens in the Baltic Sea, a species introduced historically via solid ballast in the 19th century. Although protected in Poland, the authors argue that C. connivens should not be listed as endangered in the Baltic region due to its non-native origin. Taken together, these studies underscore the multifaceted significance of Chara species—not only as ecological indicators and agents of bioremediation but also as valuable biomass resources for sustainable applications such as organic compost production. The growing body of research highlights the urgent need to reassess the ecological status, conservation priorities, and practical utility of Chara spp., particularly C. globularis, in the context of freshwater ecosystem management and restoration.
Research question: How does seasonal water quality variation influence Chara globularis biomass dynamics in Lake Titicaca, and how can sustainable harvesting be implemented to mitigate ecological impacts and promote agricultural reuse?
Specific objectives: a) To determine the physico-chemical water parameters (e.g., pH, DO, EC, TDS, temperature) that influence the biomass develop-ment of Chara globularis. b) To quantify seasonal biomass variation and assess its sustainable harvest potential for ecological control and organic fertilizer production.
METHODOLOGY
Study area
The research was carried out in Lake Titicaca, specifically in the Puno sector (Peru), at an elevation of 3810 m.a.s.l. The site experiences seasonal temperature fluctuations between 12 and 18 °C. Sampling zones were georeferenced and included shallow aquatic habitats with varying degrees of Chara coverage.
Sampling design
The study area covered 10 hectares, subdivided into two zones: Zone A (dominated by C. globularis) and Zone B (free of Chara). Monthly surveys occurred between February and July 2024. A standardized sampling ensured temporal and spatial consistency. Sampling was conducted from 07:00 to 11:00 h, coinciding with optimal environmental conditions.
Biomass quantification
A metal quadrat (0.25 × 0.25 m) mounted on a 4-meter pole was used to collect submerged vegetation. Five quadrats were randomly sampled per zone each month. Samples were gently rinsed to remove debris and weighed in the field using a digital scale (±0.01 g precision). The values obtained were extrapolated to biomass per hectare (kg/ha). These data were then scaled up to estimate the total biomass of Chara globularis across its reported distribution in Lake Titicaca, covering 43,600 hectares (Collot et al., 1983). Based on this large-scale estimate, the sustainable harvest potential was also calculated, applying a 60% extraction threshold to balance biomass utilization with ecosystem conservation.
Water quality measurements
Water physicochemical parameters (pH, DO, EC, PSU, TDS, temperature) were measured in situ using a HANNA R-2348 multiparameter probe. Triplicate readings were taken at three depths (surface, mid-water, bottom) in both zones. The data were compiled monthly to assess seasonal variation.
Nutrient composition of biomass
Collected biomass samples were air-dried, ground, and analyzed in the Environmental Chemistry Laboratory at Universidad Nacional del Altiplano. Nitrogen was determined using the Kjeldahl method, phosphorus via acid digestion and molybdenum blue spectrophotometry, and potassium by flame photo-metry. Values were expressed as percentage dry weight (% DW).
Soil analysis
Soil samples from Chara and non-Chara zones were collected and analyzed for pH, EC, total NPK, organic carbon, and organic matter. The results were evaluated through PCA to detect edaphic shifts linked to biomass presence.
Biodiversity assessment
Macrophytes and macroinvertebrates were surveyed monthly in both zones using visual identification and taxonomic keys. Species richness and seasonal occurrence were documented.
Statistical analysis
Data were analyzed using INFOSTAT software. The Kruskal-Wallis test was used to evaluate biomass variation and biodiversity across time and space. Spearman correlation and PCA were used to examine multivariate relationships between environmental parameters, biomass, and soil fertility metrics.
RESULT AND DISCUSSION
Available biomass and sustainable harvest of Chara globularis
The sustainable harvest of C. globularis is supported by its high productivity, with an estimated biomass of 24800 kg/ha covering approximately 43600 ha of the aquatic area in Lake Titicaca (Collot et al., 1983), amounting to a total biomass of over 1.08 million metric tons (1’081’280000 kg) (Table 1). If managed through a controlled partial removal, this species can be harvested without compromising the ecological balance. According to Asaeda et al. (2014), dense Chara beds can induce sediment anoxia, thereby reducing oxygen availability for other aquatic organisms. Similarly, Rodrigo et al. (2015) reported that excessive macrophyte growth can suppress plant and animal biodiversity in aquatic ecosystems. A sustainable harvest of 60%—equivalent to 648768 metric tons—enables the following benefits: a) repurposing a significant portion of biomass as organic compost, rich in carbon, nitrogen, and calcium, for the rehabilitation of degraded agricultural soils (Gette et al., 2014), b) reducing excessive plant cover, which otherwise may lead to decreased water oxygenation and promote reducing sedimentary conditions, and c) conserving the remaining 40% (432512 metric tons) of C. globularis in situ, ensuring its vital ecological functions in sediment stabilization, nutrient retention, and algal bloom control (Bakker et al., 2010).
Table 1
Estimation of biomass and sustainable harvest of Chara globularis in Lake Titicaca
|
Concept |
Value |
Unit |
|
Area covered by Chara globularis |
43600 |
ha |
|
Biomass per hectare |
24800 |
kg/ha |
|
Estimated total biomass |
1081280000 |
kg |
|
Sustainable harvest (60%) |
648768000 |
kg (648768 t) |
|
Biomass to be preserved (40%) |
432512000 |
kg (432512 t) |
Source: Adapted from Collot et al. (1983), based on a 60% sustainable harvest criterion. (Bakker et al., 2010).
Figure 1 shows the monthly variation in C. globularis biomass (kg/ha) from February to July in Lake Titicaca, Peru. Biomass values increased progressively from a minimum of 211200 kg/ha in March to a peak of 705600 kg/ha in July, indicating a strong seasonal pattern. The lowest biomass in March may be associated with environmental stressors such as lower solar radiation or hydrodynamic disturbance. From April onwards, a sustained increase is observed, reaching 476800 kg/ha in April, 492800 kg/ha in May, and 600000 kg/ha in June, suggesting favorable conditions for growth, likely due to rising temperatures and increased photosynthetic activity.
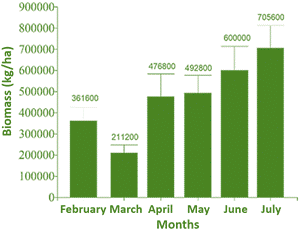
Figure 1. Estimated available biomass of Chara globularis in Lake Titicaca, Peru.
The peak in July coincides with optimal aquatic conditions for C. globularis, marking this period as the most suitable for biomass harvesting. This temporal pattern highlights the importance of aligning sustainable harvest strategies with seasonal biomass availability to maximize compost production while maintaining ecological balance.
Furthermore, the partial and strategic removal of C. globularis during its peak productivity (June) not only facilitates the exploitation of its biochemical value but also contributes to mitigating seasonal hypoxia and restoring habitat conditions for native species. This balance between conservation and productive use is essential to ensure the ecological sustainability of the lake while enhancing its role as a provider of key ecosystem services in the high Andean region.
Ecosystem impacts and habitat simplification
Table 2 reveals clear differences in macroinver-tebrate diversity between zones with and without C. globularis in Lake Titicaca. The zone without Chara shows greater taxonomic richness and temporal continuity, with five families consistently recorded across most months. In contrast, the zone with Chara exhibits reduced diversity and intermittent presence of taxa, with some species, such as Telmatobius culeus and fish eggs, appearing exclusively in this habitat. These patterns suggest that dense Chara coverage may limit macroinvertebrate diversity while providing microhabitats for specific taxa of ecological importance, highlighting the dual ecological role of C. globularis as both a potential suppressor of biodiversity and a specialized refuge.
This pattern is consistent with previous studies highlighting the critical role of alkaline conditions in promoting charophyte growth (Rodrigo et al., 2015). Elevated pH levels enhance calcification processes and photosynthetic efficiency, thereby supporting the formation of dense macrophyte stands.
Table 3 indicates a marked contrast in macroalgal diversity between zones with and without C. globularis. The zone without Chara supports a higher diversity, with up to ten families and multiple species recorded consistently across months, reflecting a more heterogeneous and favorable habitat for various aquatic plants. In contrast, the zone with Chara is largely dominated by C. globularis, with only two additional taxa (Nostoc sp. and Balliviaspongia wirrmanni) appearing sporadically. This suggests that the dense presence of C. globularis may exert a competitive exclusion effect, reducing macroalgal diversity and altering community structure.
Table 2
Diversity of macroinvertebrates in two zones (with and without presence of Chara globularis in Lake Titicaca
|
Zone without Chara globularis presence |
|||||||
|
Family |
Species |
Feb |
Mar |
Apr |
May |
Jun |
Jul |
|
Hyalellidae |
Hyalella sp. |
X |
X |
X |
X |
X |
X |
|
Glassiphoniidae |
Helobdella sp. |
X |
X |
X |
X |
X |
|
|
Cochliophidae |
Rhamphopoma sp. |
X |
X |
X |
|||
|
Planorbidae |
Taphinus sp. |
X |
X |
X |
X |
X |
|
|
Fish eggs |
X |
X |
X |
||||
|
Zone with Chara globularis presence |
|||||||
|
Hyalellidae |
Hyalella sp. |
X |
X |
X |
X |
X |
X |
|
Glassiphoniidae |
Helobdella sp. |
X |
X |
X |
X |
||
|
Telmatobiidae |
Telmatobius culeus |
X |
|||||
|
Cochliophidae |
Rhamphopoma sp. |
X |
X |
||||
|
Planorbidae |
Taphinus sp. |
X |
|||||
|
Fish eggs |
X |
X |
|||||
Table 3
Diversity of macroalgae in two zones (with and without presence of Chara globularis in Lake Titicaca
|
Zone without Chara globularis presence |
|||||||
|
Family |
Species |
Feb |
Mar |
Apr |
May |
Jun |
Jul |
|
Ruppiaceae |
Ruppia maritima |
X |
X |
X |
X |
X |
|
|
Cyperaceae |
Schoenoplectus Californicus |
X |
X |
X |
X |
X |
|
|
Haloragaceae |
Myriophyllum quitense |
X |
X |
X |
X |
X |
X |
|
Hydrocharitaceae |
Elodea potamogeton |
X |
X |
X |
X |
X |
|
|
Cladophoraceae |
Cladophora crispata |
X |
X |
||||
|
Nostocaceae |
Nostoc sp. |
X |
|||||
|
Callitricheae |
Callitriche heteropoda |
X |
|||||
|
Amblystegiaceae |
Drepanocadus sp. |
X |
X |
||||
|
Spongillidae |
Balliviaspongia wirrmanni |
X |
X |
X |
X |
||
|
Araceae |
Lemna gibba |
X |
|||||
|
Zone with Chara globularis presence |
|||||||
|
Characeae |
Chara globularis |
X |
X |
X |
X |
X |
X |
|
Nostocaceae |
Nostoc sp. |
X |
|||||
|
Spongillidae |
Balliviaspongia wirrmanni |
X |
|||||
These findings align with those of Gette et al. (2014), who linked Chara metabolic activity to ion fluxes, and Hutorowicz and Dziedzic (2008), who demonstrated that dense Chara beds can alter the ionic composition of the surrounding water column. Additionally, the reduced levels of dissolved oxygen (DO) observed in Chara-dominated zones (3.7 mg/L compared to 5.9 mg/L in non-Chara areas) support previous reports of hypoxic conditions beneath dense canopies, primarily due to restricted atmospheric exchange and elevated microbial oxygen consumption during biomass decomposition (Hutorowicz & Dziedzic, 2008).
Principal Component Analysis (PCA) further supported these relationships: biomass was negatively correlated with depth and positively associated with conductivity, salinity, and pH—factors favorable for Chara proliferation (Figure 2). Biomass accumulation during the late wet season aligns with the results of Van-Zuidam et al. (2022), who found that C. globularis maintains coverage under grazing and fluctuating physical stress. Thus, the phenology and environmental preference of C. globularis suggest that alkaline, stable, shallow habitats during June are optimal for biomass expansion and strategic harvesting.
Biofertilizer potential and circular economy
The chemical composition of C. globularis biomass demonstrates its agronomic value. The recorded potassium (30.68%), phosphorus (1.90%), and nitrogen (0.68%) levels exceed those reported in other aquatic macrophytes (Asaeda et al., 2014), and are comparable to nutrient-enriched composts. Organic matter content (17.49%) and increased sediment carbon and nutrients in Chara zones (Table 4) reveal this species’ capacity to enhance benthic fertility. These findings echo Laffont-Schwob et al. (2015), who highlighted the value of Chara in nutrient cycling and phyto-remediation.
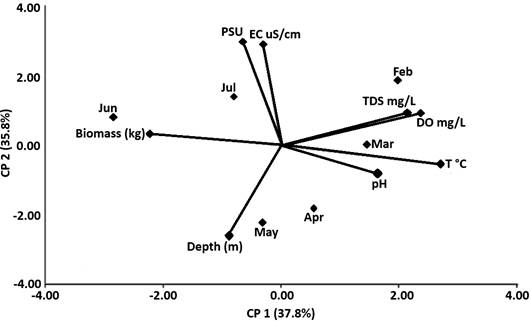
Figure 2. Principal Component Analysis of water physical parameters over time (months).
Table 4
Analysis of the physico-chemical characteristics of Chara globularis and the soil in areas with and without its presence
|
Characteristics |
Physico-chemical of Chara globularis |
Suelo Without Chara globularis |
Suelo With Chara globularis |
|
pH |
7.10 |
6.75 |
6.88 |
|
EC (mS/cm) |
1.40 |
0.98 |
1.3 |
|
Total Nitrogen (%) |
1.50 |
0.48 |
0.52 |
|
Total Phosphorus (%) |
0.68 |
0.3 |
0.33 |
|
Total Potassium (%) |
1.90 |
0.4 |
0.57 |
|
Organic Matter (%) |
30.68 |
9.6 |
10.44 |
|
Organic Carbon (%) |
17.49 |
5.57 |
6.05 |
Furthermore, Borges et al. (2023) emphasized the importance of integrating aquatic biomass into local agroecological models. Our results support this, showing that sustainable harvest of Chara biomass at its seasonal peak can contribute to organic agriculture and nutrient recycling. In addition to compost potential, Laffont-Schwob et al. (2015) and Thakur et al. (2022) noted the antimicrobial and bioenergy applications of Chara spp., the latter reporting up to 605 L CH₄/kg SV via anaerobic co-digestion. These multifaceted uses confirm the relevance of C. globularis in circular bioeconomy frameworks.
Strategic management recommendations
Zone A, dominated by Chara, exhibited reduced macrophyte and macroinvertebrate diversity compared to Zone B. This confirms the ecological homogenization described by Bakker et al. (2010), who reported that dense submerged macrophytes can monopolize light and space, suppressing other aquatic flora. The potential production of allelopathic compounds by Chara spp., as noted by Gette et al. (2014), may further limit the establishment of competing species. While overall biodiversity declined, the exclusive presence of Telmatobius culeus and Orestias eggs in Chara habitats suggests that these mats provide structural refuge for select native vertebrates, consistent with observations by Rodrigo et al. (2015). Moreover, phenological variations, such as the autumnal dominance of Lychnothamnus barbatus (Brzozowski & Pełechaty, 2024), may allow complementary species coexistence over time, reinforcing habitat resilience under climate variability. Retike et al. (2021) recommend aligning harvest windows with biomass phenology to minimize disturbance and optimize yield. Our data suggest June as the most suitable period for biomass removal, balancing peak productivity with minimal ecological impact.
Van-Onsem et al. (2018) observed stable oospore production in C. globularis despite shading from Lemna minor, supporting the species’ reproductive resilience. This is crucial for population recovery following harvest. Maredová et al. (2021) also reported limited allelopathic impact of C. globularis on cyanobacteria (15/532 assays), suggesting limited biocontrol potential but minimal risk of broader ecosystem interference. Integrating these insights with PCA findings allows adaptive management to sustain Chara populations while leveraging their ecological and productive functions.
Biogeographic context and scientific relevance
The ecological versatility of Chara spp. is further highlighted by their wide historical and biogeographical distribution. Chemeris et al. (2020) documented charophyte survival in permafrost regions, including C. globularis, due to flexible life-history traits. Similarly, Demirci et al. (2023, 2025) traced the persistence of Chara species in palaeolakes across Turkey, underlining their role in long-term aquatic succession. These findings validate our observations regarding environmental preferences and resilience of C. globularis.
Gruszka et al. (2023) discussed the debated status of Chara connivens in the Baltic Sea, proposing management decisions based on origin and ecological role. Our results echo this approach: understanding the ecological behavior of C. globularis at local scales informs broader conservation and utilization strategies. In sum, the present study builds upon and extends existing research, evidencing that C. globularis not only adapts effectively to specific limnological niches but also offers practical benefits through sustainable biomass use, reinforcing its value in ecological restoration and circular economy systems.
CONCLUSIONS
Chara globularis is estimated to cover 43,600 hectares of Lake Titicaca, with an average density of 24800 kg/ha, reaching a total biomass of 1081280000 kg. This high productivity confirms its dominance in shallow areas of the lake and highlights its potential as an abundant plant resource in high Andean aquatic ecosystems.
Based on a 60% sustainable harvest criterion, up to 648768000 kg (648768 t) of Chara globularis can be harvested without critically affecting its ecological role. This biomass could be transformed into high-value organic compost for urban green areas and degraded agricultural soils, promoting circular economy practices and local food security. Preserving the remaining 40% (432512000 kg) ensures the stability of sensitive aquatic habitats.
Areas dominated by Chara globularis showed slightly alkaline pH values (8.5 – 8.6), higher electrical conductivity (1.40 mS/cm), and lower dissolved oxygen levels (3.7 mg/L). A reduction in plant and faunal diversity was also observed in densely colonized zones. This underscores the need to manage its proliferation to prevent negative impacts on biodiversity and water quality.
REFERENCES
Asaeda, T., Senavirathna, M. D. J., Kaneko, Y., Rashid, MHV. 2014. Effect of calcium and magnesium on the growth and calcite encrustation of Chara fibrosa. Aquatic Botany, 113, 100–106. https://doi.org/10.1016/j.aquabot.2013.11.002
Bakker, E. S., Van-Donk, E., Declercq, S. A. J., Helmsing, N. R., Hidding, B., & Nolet, B. A. (2010). Effect of macrophyte community composition and nutrient enrichment on plant biomass and algal blooms. Basic and Applied Ecology, 11(5), 432–443. https://doi.org/10.1016/j.baae.2010.06.005
Borges, D., Duarte, L., Costa, I., Bio, A., Silva, J., Sousa-Pinto, I., & Gonçalves, J. A. (2023). New methodology for intertidal seaweed biomass estimation using multispectral data obtained with unoccupied aerial vehicles. Remote Sensing, 15(13), 3359. https://doi.org/10.3390/rs15133359
Brzozowski, M., & Pełechaty, M. (2024). Different seasonal dynamics of contrasting aquatic plants and the consequences for the submerged macrophyte biodiversity. Ecohydrology & Hydrobiology, 25, 225–237. https://doi.org/10.1016/j.ecohyd.2024.03.003
Collot D, Koriyama F. & García E. 1983. - Répartitions, biomasses et productions des macrophytes du lac Titicaca. Rev. Hydrobiol. trap., 16(3), 241-261.
Chemeris, E. V., Romanov, R. E., Kopyrina, L. I., Filippova, V. A., Mochalova, O. A., Efimov, D. Y., & Bobrov, A. A. (2020). How charophytes (Streptophyta, Charales) survive in severe conditions of the permafrost area in Far North-East Asia. Limnologica, 83, 125784. https://doi.org/10.1016/j.limno.2020.125784
Demirci, E., Sanjuan, J., & Tunoğlu, C. (2023). Early Pleistocene charophyte flora from Dursunlu (Ilgın Basin, Turkey): Palaeoecological implications. Review of Palaeobotany and Palynology, 311, 104848. https://doi.org/10.1016/j.revpalbo.2023.104848
Demirci, E., Sanjuan, J., Tunoğlu, C., Tuncer, A., & Bulut, Y. (2025). Late Miocene aquatic flora from the Yalvaç Basin (Central Anatolia, Türkiye): Biostratigraphy and paleoecology. Review of Palaeobotany and Palynology, 332, 105221. https://doi.org/10.1016/j.revpalbo.2024.105221
Gette-Bouvarot, M., Mermillod-Blondin, F., Lemoine, D., Delolme, C., Danjean, M., Etienne, L., & Volatier, L. (2014). The potential control of benthic biofilm growth by macrophytes: A mesocosm approach. Ecological Engineering, 75, 178–186. https://doi.org/10.1016/j.ecoleng.2014.12.000
Gruszka, P., Brzeska-Roszczyk, P., & Pełechaty, M. (2023). Alien or endangered? Historical development of Chara connivens in the Baltic coastal waters and its implication for the species management. Journal for Nature Conservation, 73, 126416. https://doi.org/10.1016/j.jnc.2023.126416
Hutorowicz, A., & Dziedzic, J. (2008). Long-term changes in macrophyte vegetation after reduction of fish stock in a shallow lake. Aquatic Botany, 88(3), 265–272. https://doi.org/10.1016/j.aquabot.2007.11.002
Laffont-Schwob, I., Triboit, F., Prudent, P., Soulié-Märsche, I., Rabier, J., Despréaux, M., & Thiéry, A. (2015). Trace metal extraction and biomass production by spontaneous vegetation in temporary Mediterranean stormwater highway retention ponds: Freshwater macroalgae (Chara spp.) vs. cattails (Typha spp.). Ecological Engineering, 81, 173–181. https://doi.org/10.1016/j.ecoleng.2015.04.052
Li, J., Wang, X., & Zhang, M. (2018). Satellite-based assessment of macroalgal biomass in freshwater ecosystems. Remote Sensing, 10(3), 498. https://doi.org/10.3390/rs10030498
Maredová, N., Altman, J., & Kaštovský, J. (2021). The effects of macrophytes on the growth of bloom-forming cyanobacteria: Systematic review and experiment. Science of the Total Environment, 792, 148413. https://doi.org/10.1016/j.scitotenv.2021.148413
Pełechaty M, Ossowska J, Pukacz A, Apolinarska K, Siepak M. 2015. Site-dependent species composition, structure, and environmental conditions of Chara tomentosa L. meadows in western Poland. Aquatic Botany, 120, 92–100. https://doi.org/10.1016/j.aquabot.2014.06.015
Retike, I., Akstinas, V., Bikše, J., Bruzgo, M., Demidko, J., Dimante-Deimantoviča, I., & Virbickas, T. (2021). Methodology of lake ecosystem health assessment. European Union under the Interreg V-A Latvia–Lithuania Programme 2014–2020.
Rybak, M., Joniak, T., Gąbka, M., & Sobczyński, T. (2017). The inhibition of growth and oospores production in Chara hispida L. as an effect of iron sulphate addition: Conclusions for the use of iron coagulants in lake restoration. Ecological Engineering, 105, 1–6. https://doi.org/10.1016/j.ecoleng.2017.04.044
Rodrigo, M. A., Rojo, C., Segura, M., Alonso-Guillén, J. L., Martín, M., & Vera, P. (2015). The role of charophytes in a Mediterranean pond created for restoration purposes. Aquatic Botany, 120, 101–111. https://doi.org/10.1016/j.aquabot.2014.05.004
Sviben, S., Matoničkin Kepčija, R., Vidaković-Cifrek, Ž., Sertić Perić, M., Kružić, P., Popijač, A., & Primc, B. (2018). Chara spp. exhibit highly heterogeneous light adaptation, calcite encrustation and epiphyton patterns in a marl lake. Aquatic Botany, 147, 1–10. https://doi.org/10.1016/j.aquabot.2018.01.007
Thakur, N., Salama, E. S., Sharma, M., Sharma, P., Sharma, D., & Li, X. (2022). Efficient utilization and management of seaweed biomass for biogas production. Materials Today Sustainability, 18, 100120. https://doi.org/10.1016/j.mtsust.2022.100120
Van-Onsem, S., Rops, J., & Triest, L. (2018). Submerged seed, turion and oospore rain: A trap quantifying propagule deposition under aquatic vegetation. Aquatic Botany, 145, 21–28. https://doi.org/10.1016/j.aquabot.2017.11.007
Van-Zuidam, B. G., Bakker, E. S., van Geest, G. J., & Peeters, E. T. H. M. (2022). Submerged vegetation colonizes behind artificial wave shelter after a 10-year time-lag and persists under high grazing pressure by waterbirds. Aquatic Botany, 181, 103541. https://doi.org/10.1016/j.aquabot.2022.103541