 |
INTRODUCCIÓN
Interacción biológica de la entomofauna en Vigna unguiculata L. asociada a la temperatura y fases fenológicas en un sistema de cultivo orgánico
Biological interaction of the entomofauna in Vigna unguiculata L. associated with temperature and phenological phases in an organic cultivation system
Eduardo Josue Villegas-Navarro1; Pedro S. Castillo-Carrillo1; César Augusto Mogollón-Farias1,2; Segundo Melecio Garcia-Garcia1,2; Cesar Alejandro Vásquez-Garcia2; Andy Josué Luna-Socola1; Jorge Luis Purizaga-Preciado1; Archi Alejandro Ruiz-Polo2; Jose Stalyn Cordova Campos1,2
1 Programa Profesional de Agronomía, Facultad de Ciencias Agrarias, Universidad Nacional de Tumbes. Ciudad Universitaria, Av. Universitaria S/N, Tumbes, Perú.
2 Estación Experimental Agraria Los Cedros. Dirección de Recursos Genéticos y Biotecnología. Instituto Nacional de Innovación Agraria (INIA), Panamericana Norte Kilómetro 12, Tumbes, Perú.
* Autor corresponsal: villegasnavarrojosue72@gmail.com (E. J. Villegas-Navarro).
ORCID de los autores:
E. J. Villegas-Navarro: https://orcid.org/0009-0001-6170-7646 A. J. Luna-Socola: https://orcid.org/0009-0003-7782-3419
P. S. Castillo-Carrillo: https://orcid.org/0000-0002-0255-1047 J. L. Purizaga-Preciado: https://orcid.org/0000-0003-1160-3715
C. A. Mogollón-Farias: https://orcid.org/0009-0000-5390-9208 A. A. Ruiz-Polo: https://orcid.org/0009-0005-1273-2625
S. M. Garcia-Garcia: https://orcid.org/0009-0003-6300-9221 J. S. Cordova-Campos: https://orcid.org/0000-0002-5891-4679
C. A. Vázques-Garcia: https://orcid.org/0009-0005-2803-7264
_______________________________________________________________________________________________________________________________
RESUMEN
Vigna unguiculata L. es un cultivo clave para la seguridad alimentaria, sin embargo, su rendimiento se ve afectado por plagas y agroquímicos que perjudican a insectos benéficos. Por ello, estudiar las asociaciones con factores bióticos y abióticos es importante, más aún en sistemas orgánicos. El presente estudio evaluó las interacciones biológicas en la entomofauna de V. unguiculata L. asociada a la temperatura y fases fenológicas en un sistema orgánico establecido en la EEA-Los Cedros del INIA-MIDAGRI (Perú), empleando semillas del cultivar vaina blanca. Se realizaron nueve evaluaciones fragmentadas en las fases vegetativa, de floración y reproductiva durante la fenología del cultivo por 45 días. El muestreo se efectuó en 20 plantas seleccionadas al azar, mediante red entomológica y aspirador manual. La identificación taxonómica se llevó a cabo con claves entomológicas, y los datos fueron analizados mediante pruebas de normalidad (Shapiro-Wilk), correlaciones de Spearman y frecuencias taxonómicas. Se registraron seis órdenes con especies como Toxomerus sp., Ocyptamus dimidiatus, Allograpta sp., Apis mellifera, Spodoptera eridania (polinizador), Zelus sp., Hippodamia convergens, Cycloneda sanguinea, Chrysoperla externa (depredador), especie de la familia Lygaeidae, Sibovia sp., Empoasca kraemeri (plaga), Rupela albina (migratorio y/o visitante), Digonogastra sp. (Parasitoide). Se observó que la entomofauna varió por etapa fenológica del cultivo, siendo Hymenoptera el orden más abundante, seguido de Hemiptera, Diptera y Coleoptera, mientras que Lepidoptera y Neuroptera fueron menos frecuentes. La temperatura promedio fue de 28,2 °C, sin correlaciones significativas con la abundancia de insectos. Se infiere que V. unguiculata L. manejado con un sistema orgánico favorece diversas interacciones biológicas, donde su fenología es un factor determinante en la dinámica de las especies de insectos, mostrando una influencia más significativa que la propia temperatura.
Palabras clave: Biología; Entomología; Fenología; Insectos; Relaciones interespecíficas.
ABSTRACT
Vigna unguiculata L. is a key crop for food security; however, its yield is affected by pests and agrochemicals that harm beneficial insects. Therefore, studying its associations with biotic and abiotic factors is important, especially in organic systems. The present study evaluated the biological interactions within the entomofauna of V. unguiculata L. in relation to temperature and phenological stages in an organic system established at the EEA–Los Cedros of INIA–MIDAGRI (Peru), using seeds of the white pod cultivar. Nine assessments were conducted across the vegetative, flowering, and reproductive stages during the crop’s phenology over a period of 45 days. Sampling was carried out on 20 randomly selected plants using an entomological net and a manual aspirator. Taxonomic identification was performed using entomological keys, and the data were analyzed through normality tests (Shapiro–Wilk), Spearman correlations, and taxonomic frequencies. Six orders were recorded, including species such as Toxomerus sp., Ocyptamus dimidiatus, Allograpta sp., Apis mellifera, Spodoptera eridania (pollinator), Zelus sp., Hippodamia convergens, Cycloneda sanguinea, Chrysoperla externa (predator), a species from the family Lygaeidae, Sibovia sp., Empoasca kraemeri (pest), Rupela albina (migratory and/or visitor), and Digonogastra sp. (parasitoid). It was observed that the entomofauna varied according to the crop’s phenological stage, with Hymenoptera being the most abundant order, followed by Hemiptera, Diptera, and Coleoptera, while Lepidoptera and Neuroptera were less frequent. The average temperature was 28.2 °C, with no significant correlations with insect abundance. It is inferred that V. unguiculata L. managed under an organic system promotes diverse biological interactions, where phenology is a determining factor in insect species dynamics, showing a more significant influence than temperature itself.
Keywords: Biology; Entomology; Phenology; Insects; Interspecific Relationships.
________________________________________________________________________________________________________________________________________________________
Recibido: 16-09-2025.
Aceptado: 29-03-2026.
|
Vigna unguiculata L., conocido comúnmente como frijol caupí o frijol de carita, es una leguminosa de gran importancia agrícola y nutricional en regiones tropicales y subtropicales, debido a su alto contenido proteico, su capacidad de fijación biológica de nitrógeno y su adaptabilidad a diferentes condiciones edafoclimáticas (Gonçalves et al., 2016; Abebe & Alemayehu, 2022). Sin embargo, su rendimiento generalmente suele ser bajo debido a la actividad de diversas plagas que afectan el cultivo en el campo en diferentes etapas de su fenología (Caldera & Jarquín, 2024; Ansar et al., 2024).
A nivel mundial, las principales plagas que afectan gravemente al caupí durante todas sus etapas de desarrollo son el barrenador de vainas (Maruca vitrata Fabricius), los trips de yemas florales (Megalurothrips sjostedti Trybom), los escarabajos del follaje (Ootheca sp., Medythia sp.), el pulgón del caupí (Aphis craccivora Koch) y un complejo de chinches chupadoras, entre las que destacan Clavigralla spp., Anoplocnemis sp., Riptortus sp., Mirperus sp., Nezara viridula Fab. y Aspavia armigera L. Sin un manejo adecuado, estas plagas reducen significativamente el rendimiento del grano (Jackai y Daoust, 1986). Desde hace décadas hasta la actualidad se han propuesto diversas estrategias de control, los insecticidas químicos siguen siendo los más eficaces y permiten incrementos notables en la producción (Carter et al., 1989; Mateos et al., 2025).
No obstante, estas plagas pueden variar según la ubicación geográfica. En ese sentido, a nivel local, en Perú, entre las especies descritas como plagas del frijol caupi (V. unguiculata L.), se encuentran los “gusanos cortadores” o “gusanos de tierra” Agrotis ipsilon y Spodoptera eridania (Noctuidae), que dañan plántulas al cortar tallos y hojas; el “gusano picador del tallo” Elasmopalpus lignosellus (Pyralidae), que barrenan tallos jóvenes; el “gusano pegador de hojas” Omiodes indicata (Crambidae), que consume el parénquima foliar; el “escarabajo de hojas” o “mosquilla” Cerotoma fascialis (Chrysomelidae), que perfora hojas, flores y vainas; la “cigarrita verde” Empoasca kraemeri (Cicadellidae), que succiona savia y causa deformaciones foliares; el “pulgón de la melaza” Aphis craccivora (Aphididae), que reduce el vigor de la planta; la “mosca blanca” Bemisia tabaci (Aleyrodidae), vector de virosis; y las “moscas minadoras” Liriomyza sativae, L. huidobrensis y L. quadrata (Agromyzidae), cuyas larvas minan las hojas disminuyendo la fotosíntesis. Además, el “gusano perforador de vainas” Cydia fabivora (Tortricidae) destruye semillas dentro de las vainas, mientras que el “picudo de las vainas” Chalcodermus aeneus (Curculionidae) daña granos y vainas en desarrollo (Castillo, 2013).
Frente a esta problemática, la aplicación de insecticidas se ha consolidado como una estrategia utilizada desde hace varias décadas en este cultivo (de Freitas Bueno et al., 2011), así como en otros de gran relevancia agrícola (Urías-Rivas et al., 2024; Ruiz-Sánchez et al., 2025).
No obstante, desde hace muchos años se reconoce que la aplicación de insecticidas en agroecosiste-mas altera la biodiversidad, siendo los artrópodos los más afectados, generando un aumento en la abundancia de plagas (resurgimiento) y la apari-ción de nuevas especies plaga, como consecuencia de la eliminación de sus depredadores y paras-itoides naturales. Asimismo, este efecto, derivado de la acción directa de los insecticidas sobre organismos no objetivo, ha sido documentado de manera consistente en ensayos de campo y en estudios a gran escala con insecticidas de amplio espectro (Metcalf, 1980; Godfray & Chan, 1990; Mochizuki, 2003).
La interacción entre V. unguiculata L. y la entomo-fauna asociada constituye un componente esencial que debe estudiarse en su agroecosistema, dado que distintos grupos de insectos cumplen fun-ciones ecológicas clave como polinizadores, herbí-voros, depredadores y parasitoides, regulando directa o indirectamente la dinámica de la entomofauna en el cultivo (Jankielsohn, 2018; Sahani et al., 2023; Jasrotia et al., 2023). Por ello, los sistemas de producción orgánica surgen como una alternativa sostenible, favoreciendo la diversifica-ción agrícola, la seguridad alimentaria y la dismi-nución en el uso de insumos químicos sintéticos (Sharma et al., 2024; Da Costa et al., 2024).
En ese sentido, los factores abióticos, como la temperatura, son objeto de estudio puesto que influyen de forma determinante en la abundancia, composición y actividad de las comunidades de insectos (Baltazar et al., 2016; Ruiz y Barrera, 2024), siendo el verano la temporada en la que se manifiesta con más intensidad en zonas tropicales, lo cual causa junto con la humedad un incremento en las interacciones biológicas entre distintos organismos en diversos cultivos agrícolas. (Skendžić, 2021; Subedi et al., 2023). lo cual es importante comprender, ya que, al determinar la diversidad entomológica en función del clima, es posible entender y establecer estrategias de manejo integrado de plagas y conservación de organismos benéficos en sistemas productivos agrícolas (Macchia, 2024; Baltazar, 2016).
En virtud de lo anteriormente descrito, el estudio de las interacciones biológicas en la entomofauna de V. unguiculata L. es esencial para comprender cómo factores bióticos y abióticos influyen en la dinámica de los insectos asociados a este cultivo clave para la seguridad alimentaria. Asimismo, evaluar la relación entre la temperatura y las fases fenológicas en un sistema orgánico permite identificar patrones de abundancia y diversidad de insectos, incluyendo polinizadores, depredadores y plagas, sin la interferencia de agroquímicos. Dicha información es fundamental para optimizar el manejo agronómico sostenible, favorecer servicios ecosistémicos y mitigar el impacto de plagas, contribuyendo al fortalecimiento de prácticas agrícolas que promuevan la biodiversidad y la productividad en sistemas orgánicos.
El objetivo del presente estudio fue evaluar las interacciones biológicas en la entomofauna de V. unguiculata L. asociada a la temperatura y fases fenológicas en un sistema de cultivo orgánico.
METODOLOGÍA
Diseño y área del estudio
El estudio posee un diseño no experimental, enfoque cuantitativo, nivel descriptivo tipo transversal, el cual se llevó a cabo entre el 26 de octubre y 28 de diciembre del 2024 en la estación experimental agraria (EEA) Los Cedros del Instituto Nacional de Innovación Agraria (INIA) del Ministerio de Desarrollo Agrario y Riego (MIDAGRI), ubicada al norte de Perú, en el caserío Los Cedros en el departamento de Tumbes (Figura 1). El clima se caracteriza por una temperatura promedio de 22,2 °C, alcanzando valores medios máximos de 29,5 °C y mínimos de 22,9 °C; asi-mismo, presenta una humedad relativa promedio de 88,1%, con máximos de 90% y mínimos de 85,2% (Servicio Nacional de Meteorología e Hidrología del Perú [SENAMHI], 2025).
Instalación en campo del cultivo V. unguiculata L
La instalación del cultivo se llevó a cabo en cinco etapas: (1) limpieza del terreno, (2) labores de labranza, (3) trazado de surcos, (4) riego inicial y (5) siembra. El área de estudio comprendió 200 m², con un suelo de textura franco arcilloso-arenosa, aislado de otros cultivos presentes en la EEA Los Cedros-INIA. El material genético empleado correspondió a semillas de caupí, proporcionadas por la EEA Los Cedros, con las siguientes especificaciones técnicas:
Especie: Vigna unguiculata (caupí).
Cultivar: Vaina blanca – INIA.
Lote N.°: TUM-01-2024-1.
Productor: INIA – EEA Los Cedros.
Lugar de producción: Corrales, Tumbes.
Manejo del cultivo
Se espolvoreó azufre en presentación de polvo seco (PD) a una dosis de 18 kg/ha como un controlador acaricida (contra ácaros) y fungicida (contra hongos como el oídio). El control de malezas se efectuó manualmente. Durante el periodo de evaluación del cultivo se realizaron tres riegos en un lapso de 40 días cada 10 días, complementados con labores de aporque para favorecer el desarrollo de las plantas.
Evaluación y recolección de insectos
Las evaluaciones se centraron en la recolección de insectos adultos durante las fases vegetativa, de floración y reproductiva del cultivo, efectuándose cada tres días a lo largo de un periodo de 45 días.
En total se realizaron nueve muestreos: tres distribuidos entre octubre y noviembre, tres en noviembre y tres entre noviembre y diciembre. Las restricciones de tiempo limitaron la identificación de un mayor número de insectos, lo que afectó el material biológico disponible y redujo la posibilidad de ampliar el número de evaluaciones.
Figura 1. Mapa de la ubicación geográfica del área en la que se ejecutó el estudio.
El método de captura empleado consistió en la selección de 20 plantas de forma aleatoria, seguido de un muestreo utilizando dos técnicas: (I) redada doble de forma lineal mediante una red entomológica de 40 cm de diámetro x 70 cm de alto para insectos de mayor tamaño y (II) succión empleando un aspirador entomológico manual para insectos de menor tamaño (Lima et al., 2024).
Los registros se consignaron en fichas de evaluación, anotando tanto la abundancia de individuos recolectados como las fechas de observación. Posteriormente, los especímenes fueron acondicionados en bolsas plásticas, placas Petri o frascos de vidrio y trasladados a las instalaciones del área de montaje de insectos del museo entomológico de la Universidad Nacional de Tumbes (3°34′58″S 80°26′39″O) donde se llevó a cabo la identificación taxonómica.
Identificación taxonómica
Los insectos recolectados se identificaron a nivel de orden y especie empleando pinceles N° 00, pinzas entomológicas y guías taxonómicas descritas por Castillo (2013) y Ferrufino (2010).
Evaluación de temperatura
La temperatura se registró empleando un termómetro digital de ambiente modelo Clock & Hygo modelo BOECO Germany. Las temperaturas se registraron y cotejaron con las disponibles para la zona en la base de datos del Servicio Nacional de Meteorología e Hidrología del Perú.
Análisis de datos
Los datos recolectados se transfirieron a matrices en hojas de cálculo del software Microsoft Excel v.16, con el que se analizó la frecuencia de las poblaciones de insectos, seguido de análisis en el software R Studio (v.4.5), aplicando inicialmente la prueba de Shapiro-Wilk con el fin de verificar la normalidad de la distribución de los datos (p-valor > α = 0.05) o su desviación de la misma (p-valor ≤ α = 0.05), lo que permitió seleccionar el coeficiente de correlación apropiado (Spearman o de Pearson). La aplicación de Spearman se realizó con los paquetes Hmisc para el cálculo de la matriz de correlación, y corrplot junto con ggcorrplot para la visualización gráfica de los coeficientes de correlación, y ggplot2 para personalizar la escala de colores.
Aspectos éticos
La investigación fue autorizada por la Facultad de Ciencias Agrarias de la Universidad Nacional de Tumbes mediante el Memorándum N.º 013-2023/UNTUMBES-VRACAD-FCA–PAA. Asimismo, contó con el permiso del Instituto Nacional de Innovación Agraria (INIA) del Ministerio de Desarrollo Agrario y Riego (MIDAGRI) del Perú a través del Oficio N.º 0036-2024-MIDAGRI-INIA-UO.EEA.LC-T/D.
RESULTADOS Y DISCUSIÓN
Identificación taxonómica e interacciones biológicas
En el cultivo de V. unguiculata L. se identificaron insectos pertenecientes a distintos órdenes, entre depredadores, parasitoides, plagas, visitantes y/o migratorios y polinizadores (Figura 2), lo que evidencia la presencia de diversas interacciones biológicas en dicho agroecosistema.
De acuerdo con Saleem et al. (2024), las relaciones entre insectos y vegetales desempeñan un papel vital en el mantenimiento del equilibrio ecológico y la mejora de la productividad agrícola. En ese sentido, Dewi et al. (2020) señalan que, en las comunidades de artrópodos se pueden hallar interacciones biológicas en las que las especies cumplen roles como plagas, controladores biológicos, polinizadores, descomponedores o parasitosides. Esto ha sido comprobado con estudios basados en las interacciones entre el caupí (V. unguiculata L.) y los insectos polinizadores (Lazaridi et al., 2023). Sin embargo, Outhwaite et al. (2022), señalan que el equilibrio entre las comunidades de estos artrópodos puede verse afectado por factores bióticos y abióticos, como el cambio climático, el cual genera variaciones en la composición y el reemplazo de especie.
Dentro del orden Coleoptera se hallaron especies (Coccinellidae: Hippodamia convergens, Cycloneda sanguinea), que se caracterizan por alimentarse de otros insectos. Giorgi et al. (2009), señalan que la familia Coccinellidae comprende escarabajos de hábitos que consumen diversas familias de insectos, aunque igualmente son capaces de aprovechar recursos tróficos de origen vegetal, como el follaje. Estudios experimentales basados en la depredación, reproducción y desarrollo de la catarina H. convergens, demuestran que esta especie, en su estado larvario se alimentan de áfidos, mientras que, en la adultez, de áfidos, espigas de maíz y plantas no cultivadas (Naranjo et al., 2022). En ese contexto, la presencia de H. convergens en V. unguiculata L puede explicarse debido a su hábito alimenticio de adulto. Esto es respaldado por Funichello et al. (2012), quienes evidenciaron qué, C. sanguinea es un depredador eficiente de áfidos. Lo cual explicaría el hallazgo de C. sanguinea, ya que también fue una de las especies identificadas en el cultivo de frijol caupí evaluado.
En lo que corresponde al orden Díptera, se registraron especies (Syrphidae: Toxomerus spp., Ocyptamus dimidiatus, Allograpta sp.) con un rol benéfico, ya que sus larvas actúan como depredadores de plagas, mientras que los adultos cumplen funciones de polinizadores. Según Lucas et al. (2020), especies del género Toxomerus, como T. duplicatus muestran asociaciones directas con plagas como Myzus persicae en lechuga y Brevicoryne brassicae en brócoli, contribuyendo al control biológico en sistemas hortícolas orgánicos. De acuerdo con Hopper et al. (2011), especies de sirfidos como T. marginatus, se caracteriza por ser depredadora, reforzando la importancia de este grupo de insectos como agentes de regulación natural de poblaciones de insectos plaga.
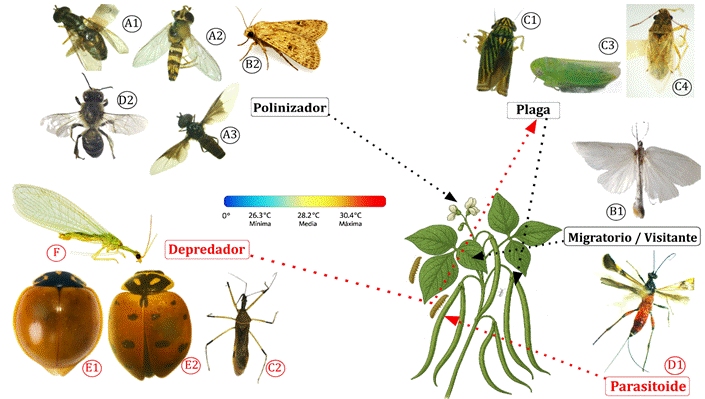
Figura 2. Especies de insectos halladas en el frijol caupí “Vigna unguiculata L.” var. vaina blanca - INIA. (A1) Toxomerus sp... (A2) Allograpta sp.. (A3) Ocyptamus dimidiatus. (B1) Rupela albina. (B2) Spodoptera eridania. (C1) Sibovia sp. (C2) Zelus sp. (C3) Empoasca kraemeri. (C4) Especie de la familia “Lygaeidae”. (D1) Digonogastra sp. (D2) Apis mellifera L. (E1) Cycloneda sanguinea. (E2) Hippodamia convergens. (F) Chrysoperla externa.
Nota. Basado en estudios previos sobre los roles biológicos de la entomofauna, la figura ilustra las relaciones interespecíficas entre las especies adultas presentes en el agroecosistema de frijol caupí V. unguiculata L. var. vaina blanca–INIA. Las flechas, recuadros y términos resaltados en rojo señalan especies clave responsables de mantener el equilibrio ecológico del sistema. La presencia de Digonogastra sp. habría estado influenciada por larvas de huéspedes. La escala de la temperatura indica valores mínimos, medios y máximos registrados durante el estudio. Los órdenes taxonómicos se distinguen mediante letras mayúsculas: (A) Diptera, (B) Lepidoptera, (C) Hemiptera, (D) Hymenoptera, (E) Coleoptera, (F) Neuroptera.
Esto coincide con reportes de Carabalí et al. (2018) y Alfonso et al. (2024), quienes señalan que, los sírfidos en su etapa adulta son visitantes frecuentes de las flores. Este hallazgo resulta relevante porque constituye el primer registro de su asociación con V. unguiculata L., un cultivo donde su presencia no había sido reportada previamente. De acuerdo con Hopper et al. (2011) y Sturza et al. (2014), hasta ahora, estos insectos habían sido vinculados a otras especies vegetales. Sin embargo, el haber hallado a Toxomerus spp. en su fase adulta, el rol que habría cumplido en el agroecosistema del frijol caupí sería el de ser polinizador.
En relación con O. dimidiatus y Allograpta sp. (Diptera: Syrphidae), diversos estudios resaltan su relevancia ecológica y agrícola, lo cual podría explicar su presencia en el agroecosistema del frijol caupí. Según Arcaya et al. (2017), Allograpta exotica constituye un importante enemigo natural de plagas comunes, particularmente de los áfidos (Hemiptera), entre ellos Aphis craccivora, reconocido como una de las principales plagas del caupí (V. unguiculata L.) y de otras leguminosas. Asimismo, estudios tanto en condiciones de laboratorio como de campo han documentado las tasas de depredación, el desarrollo larvario y el potencial controlador de especies del género Allograpta, especialmente A. exotica y A. obliqua, sobre distintos áfidos como Chaetosiphon spp. (Arcaya et al., 2017; Díaz et al., 2024). En este sentido, la presencia de Allograpta sp. en Vigna unguiculata puede asociarse a la disponibilidad de insectos hemípteros observados en el presente estudio.
Por otra parte, el registro de O. dimidiatus se explica debido a la amplitud de la distribución del género en la región neotropical. Además, de que sus adultos se caracterizan por visitar flores, donde cumplen el rol de polinizar especies de plantas silvestres y cultivadas (Doyle et al., 2020). Sin embargo, se ha documentado que otras especies del mismo género, como O. gastrostactus, presentan un notable potencial como agentes de control biológico, dado que en su fase larvaria manifiestan comportamiento depredador sobre áfidos como Sarucallis kahawaluokalani (Murgas, 2024).
En este estudio también se registró la presencia del orden Lepidóptera representado por la especie R. albina y S. eridania. En cuanto a R. albina, esta especie se caracteriza por un hábito alimenticio fitofago, siendo la principal plaga de los cultivos de arroz (Coloma et al., 2023). Estudios previos realizados en sistemas arroceros de Ecuador reportaron a esta especie junto con otras entre las principales plagas que afectan al cultivo de arroz (Sánchez et al., 2023). Por tanto, la identificación de R. albina en V. unguiculata L. se debe a la proximidad de amplias zonas arroceras en el área de estudio (INIA-EEA Los Cedros), lo que habría favorecido su desplazamiento hacia este cultivo. Esto es respaldado por la capacidad migratoria de la especie durante periodos de clima seco (Vivas y Astudillo, 2022; Horgan, 2023). Esto coincide con las características ambientales del departamento de Tumbes, el cual se define por un clima cálido y árido, con una temperatura promedio anual de 22,2 °C, alcanzando máximas de 29,5 °C y mínimas de 22,9 °C (Plataforma del Estado Peruano, s.f.; SENAMHI, 2025), lo cual se vio en la temperatura medida al momento del estudio (28,2 °C con valores entre 26,3 y 30,4 °C). Por tanto, la presencia R. albina en V. unguiculata L. se define como un visitante o insecto migratorio, más no como una plaga. Por otra parte, la presencia de S. eridania en el frijol caupí podría explicarse debido a que, en su etapa larvaria, es una plaga ya registrada para el frijol caupí (Lobato et al., 2015).
En cuanto al orden Hymenóptera, se registró la presencia de Digonogastra spp., la cual desempeña un papel significativo como parasitoide en la regulación natural de Diatraea grandiosella en México (Ramos et al., 2023). Estudios previos han documentado la ocurrencia de este género en diversos cultivos, por ejemplo, en yuca, donde fue identificado como una avispa parasitoide (Álamo et al., 2024). De igual manera, Valente et al. (2018) reportaron por primera vez en Brasil que Digonogastra spp. actúa como parasitoide de larvas de D. flavipennella. De forma análoga, investigaciones realizadas en México confirmaron su función como insecto parasitoide en Monoctenus sanchezi (Salas et al., 2023). En este contexto, la detección de Digonogastra spp. en V. unguiculata L. podría atribuirse a su estrategia de búsqueda de hospederos, ya que estos parasitoides se desplazan hacia los cultivos donde se encuentran sus presas. No obstante, dentro del mismo orden también se identificó la especie A. mellifera L., cuya presencia en el cultivo de frijol caupí se asocia a su comportamiento trófico, desempeñando funciones de polinización. Según Khalifa et al. (2021), las abejas melíferas (A. mellifera) representan un componente fundamental para el equilibrio de los ecosistemas y la producción agrícola mundial, principalmente por los servicios de polinización que proporcionan. En ese contexto, en el Perú, esta especie ha sido registrada en 17 apiarios localizados en la costa norte, abarcando los departamentos de Tumbes, Piura, Lambayeque y La Libertad (Díaz et al., 2021). En consecuencia, su detección en frijol caupí resulta comprensible.
Por otro lado, se identificaron insectos del orden Hemíptera que desempeñan distintos roles ecológicos dentro del agroecosistema. Entre ellos, se registraron ejemplares de la familia Lygaeidae, cuya identificación no pudo precisarse a nivel de especie, pero cuyo registro resulta relevante para este estudio debido a su carácter fitófago. También se detectó E. kraemeri (Cicadellidae), considerada una plaga clave en cultivos de leguminosas. De igual manera, se halló Zelus spp. (Reduviidae), reconocido por su comportamiento depredador, y Sibovia sp. (Cicadellidae), una especie conocida por actuar como vector de fitopatógenos. La presencia de estos grupos puede explicarse por las interacciones biológicas que se establecen entre insectos plaga y enemigos naturales dentro de las comunidades de artrópodos (Dewi et al., 2020). En ese sentido, estudios previos en frijol caupí (V. unguiculata L.) ya han reportado la presencia de Zelus sp. como agente de control natural contra plagas (Leite et al., 2011), lo cual coincide con lo descrito por Liccardo et al. (2020), quienes destacan el papel de Z. renardii en la depredación de Philaenus spumarius, un insecto chupador de savia de relevancia agrícola. Asimismo, el hallazgo de Sibovia sp. también encuentra respaldo en lo señalado por Moya et al. (2025), quienes han reportado su alimentación a partir del xilema en leguminosas como Acacia pennatula, mientras que Vaidya et al. (2017) reconocen a Sibovia spp. como un áfido que afecta a distintos cultivos agrícolas. Por su parte, la presencia de E. kraemeri en frijol caupí, se explica debido a que es una plaga de dicho cultivo (González et al., 2016), que viene siendo reportada desde años atrás (Díaz et al., 2025). No obstante, en otras partes del mundo como México también se ha reportado como una plaga del frijol caupí (Rojas et al., 2025. Por tanto, los resultados evidencian que en el agroecosistema coexisten hemípteros plaga y benéficos, lo que resalta la necesidad de continuar investigando estas interacciones para un manejo sostenible del cultivo.
Respecto al orden Neuróptera, se halló la presencia de C. externa asociada al cultivo de frijol caupí, lo que respalda su amplia distribución y adaptación a distintos sistemas agrícolas. Puesto que, inves-tigaciones previas han demostrado que las larvas de tercer estadio de C. externa ejercen un papel depredador efectivo sobre poblaciones de Macrosiphum euphorbiae en cultivos de rosa en Brasil (Gamboa et al., 2016). De manera similar, en Chile se han registrado C. asoralis y C. argentina como depredadores asociados a cítricos (Reguilón et al., 2023), mientras que en cultivos de maracuyá (Passiflora edulis), se ha identificado a C. externa junto a Neohydatothrips signifer como controla-dores naturales de trips (Bastidas et al., 2010).
No obstante, en su etapa adulta estas especies no mantienen hábitos depredadores, sino que contribuyen a la polinización al alimentarse principalmente de néctar y polen (Andrade et al., 2017). En el contexto local del presente estudio, la presencia de C. externa en el departamento de Tumbes ya ha sido reportada anteriormente, pero vinculada a cultivos de limón (Citrus aurantifolia) en zonas citrícolas como Matapalo, La Palma, el Centro Experimental Los Tumpis, Francos, Vaquería y Oidor entre 2012 y 2013 (Cedillo et al., 2020). Por lo tanto, la detección de C. externa, un insecto perteneciente a un género ampliamente reconocido por su actividad depredadora en etapa larvaria y polinizadora en etapa adulta podría explicarse por su elevada capacidad de interacción con cultivos agrícolas. Asimismo, es probable que esta especie desempeñe un rol ecológico dual en los cultivos de frijol caupí, actuando tanto como depredador en sus fases larvarias como polinizador en su etapa adulta. No obstante, esta hipótesis debe considerarse con cautela, ya que en el presente estudio solo se registraron individuos adultos, lo que sugiere que, en este caso particular, solo para este estudio, su función principal habría sido la de polinizador.
En síntesis, los resultados obtenidos y la infor-mación contrastada muestran la existencia de un equilibrio entre insectos fitófagos y sus enemigos naturales, lo cual indica la estabilidad del cultivo. Por ello, el manejo orgánico surge como una estrategia clave para conservar la biodiversidad funcional en el frijol caupí (V. unguiculata L.).
Entomofauna asociada a la temperatura
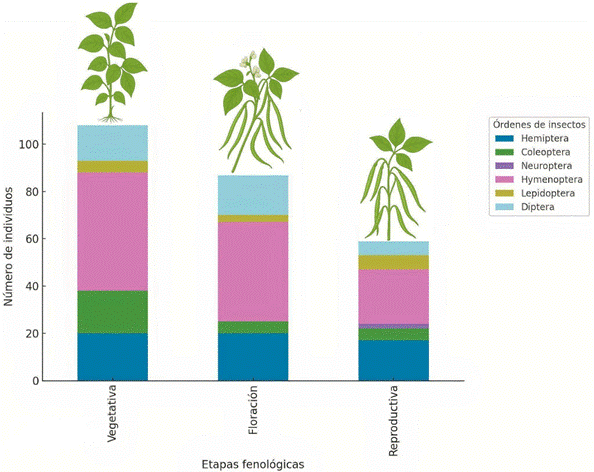
El análisis descriptivo (Tabla 1), muestra que la temperatura registró un promedio de 28,2 °C (±1,42), con valores entre 26,3 y 30,4 °C, distribuyéndose de manera normal (p = 0,660). Entre los insectos, el orden más abundante fue Hymenóptera con una media de 12,1 individuos (±7,83), seguido de Hemíptera (6,33 ± 6,08), Díptera (4,33 ± 4,42) y Coleóptera (3,33 ± 2,29), mientras que Lepidóptera y Neuróptera presen-taron las menores abundancias (2,00 ± 1,80 y 0,67 ± 1,41, respectivamente).
En cuanto a la normalidad de los datos, solo Neuróptera presentó una distribución no normal (p < 0,001), mientras que los demás órdenes y la temperatura cumplieron con este supuesto (p > 0,05). Estudios previos de Valarezo et al. (2022) aplicaron la prueba de Shapiro para comprobar la normalidad de los datos al evaluar la diversidad de insectos en Citrus aurantifolia y en la vegetación circundante de dos localidades ecológicas en Ecuador. Asimismo, otros estudios de correlación entomológica también han empleado esta prueba para determinar la distribución normal o no de sus datos, evidenciando que el resultado depende principalmente del tamaño de la muestra y del valor p obtenido (Doria et al., 2021; Nadal et al., 2024; Santos et al., 2024). En este contexto, la distribución normal observada en los datos entomológicos (órdenes de insectos) y abióticos (temperatura) puede atribuirse al tamaño amplio de las muestras en cada caso. No obstante, la distribución no normal registrada para Neuróptera podría deberse al número reducido de especies en su submuestra. Por lo tanto, para estudios de correlación estadística entre factores abióticos y bióticos, como el análisis de la entomofauna en relación con la temperatura resulta esencial asegurar un tamaño de muestra representativo, información relevante a considerar. Sin embargo, es importante señalar que la variabilidad de una muestra en estudios que incluyan organismos vivos como los insectos, no se puede controlar (evidenciado con la cantidad de especies del orden Neuróptera) puesto que no son variables que se puedan manipular, lo cual resalta la importancia de investigaciones como esta, ya que al entender mejor las interacciones biológicas y el grado en qué se manifiestan, contribuirán al diseño y planeamiento de estrategias para una agricultura sustentable y/o sostenible.
Por otro lado, el análisis de correlaciones de Spearman muestra que la temperatura no presentó correlaciones significativas con la abundancia de los órdenes de insectos evaluados (p > 0,05 en todos los casos) (Figura 4).
Tabla 1
Análisis de estadística descriptiva y prueba de normalidad de Shapiro-Wilk
|
Descriptivas |
Temperatura |
Hymenoptera |
Coleoptera |
Neuroptera |
Hymenoptera |
Lepidoptera |
Diptera |
|
Media |
28,2 |
6,33 |
3,33 |
0,667 |
12,1 |
2,00 |
4,33 |
|
Mínimo |
26,3 |
0,00 |
1,00 |
0,00 |
3,00 |
0,00 |
0,00 |
|
Máximo |
30,4 |
15,0 |
8,00 |
4,00 |
22,0 |
5,00 |
12,0 |
|
W de Shapiro-Wilk |
0,947 |
0,862 |
0,831 |
0,564 |
0,848 |
0,899 |
0,890 |
|
Valor p de Shapiro-Wilk |
0,660 |
0,100 |
0,046 |
<,001 |
0.070 |
0.247 |
0,198 |
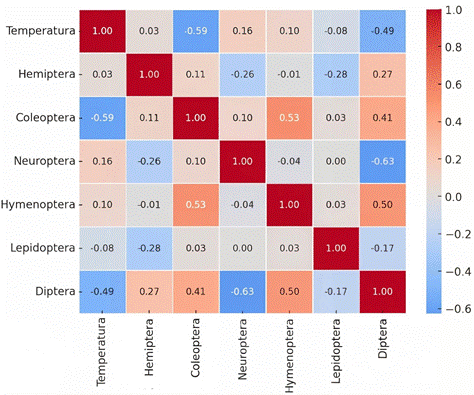
Figura 4. Mapa de calor de la matriz de correlación de la temperatura ambiental y los órdenes de insectos identificados.
Como se observa en la Figura 4, el coeficiente de Spearman halló una correlación negativa moderada con Coleóptera (rho = –0,586; p = 0,098) y Díptera (rho = –0,494; p = 0,177), lo que sugiere una tendencia a que el incremento de la temperatura se asocie con una menor presencia de estos órdenes, aunque sin alcanzar significancia estadística. En cambio, con Hemíptera (rho = 0,034), Hymenóptera (rho = 0,102), Lepidóptera (rho = –0,078) y Neuróptera (rho = 0,160) las correlaciones fueron muy bajas y sin relevancia biológica. En conjunto, los resultados indican que la temperatura no fue un factor determinante en la variación de los órdenes de insectos registrados en este estudio. Autores como Bazán & Anthony (2016), señalan que la temperatura tiene un efecto directo en los insectos. Esto es respaldado por estudios realizados décadas atrás, en los que se ha demostrado el efecto de la temperatura sobre el ciclo biológico de insectos como Lucilia sericata (Grassberger & Reiter, 2001), y estudios recientes en los que se ha evidenciado que, el dimorfismo sexual en insectos como Aedes aegypti, también se ve influenciado por la temperatura (Ruiz & Barrera, 2024).
Asimismo, estudios de Cabrera et al. (2021), han evidenciado que factores abióticos como la temperatura ambiental, temperatura del suelo y la humedad poseen una correlación con la composición de las familias de Hymenóptera, afectando la abundancia y composición de estos insectos. De manera similar, estudios en Spartium junceum L. (Fabaceae), especie presente en el valle del Mantaro de Perú, evidenciaron una alta diversidad de insectos (10 órdenes, 32 familias, 35 géneros y 16 especies), cuya composición se vio influenciada por factores climáticos, registrándose una reducción en la diversidad y heterogeneidad con el aumento de la precipitación y la humedad relativa, así como efectos adicionales por la disminución de las horas de sol (Baltazar, 2016).
En contraste, resultados opuestos se han hallado en estudios previos. Nehybová & Horak (2024), quienes han estudiado la diversidad de insectos polinizadores y su respuesta a recursos florales, y factores abióticos (temperatura, humedad, precipitación y vientos), han hallado que la diversidad de especies no se asoció con la temperatura, humedad y viento, pero sí con el proceso de floración y precipitación. Esta discordancia entre la relación significativa o no de la temperatura y la entomofauna puede explicarse debido a que actualmente el planeta enfrenta un cambio climático marcado (Vidal et al., 2025). Según Schneider et al. (2022), el impacto del cambio climático, en la relación entre plagas de insectos y cultivos, no es uniforme, sino que varía según la variable climática involucrada (tempe-ratura, sequía, precipitaciones, CO₂, etc.) y la región geográfica donde ocurre la interacción. Esto es consistente con estudios en los que se ha observado respuestas complejas insectos plagas frente al calentamiento global (Lehmann et al., 2020).
En relación con lo anteriormente descrito, los resultados obtenidos, se interpretan a modo de que la fenología del frijol caupí (V. unguiculata L.) desempeña un papel más decisivo que la temperatura en la conformación de la entomofauna a nivel de orden, lo cual se evidenció al comparar las fases vegetativa, de fructificación y reproductiva del cultivo (Figura 3). Este patrón resulta coherente al considerar que las interacciones entre factores abióticos y comunidades de insectos deben ser comprendidas en un escenario ecológico específico, dado que los efectos pueden variar según el ambiente. Asimismo, la corta duración de las etapas fenológicas de V. unguiculata L. favorece una sucesión entomológica acelerada, lo cual también explicaría la falta de correlación entre los datos. En ese sentido, se puede inferir la necesidad de replicar este estudio considerando un mayor número de evaluaciones de entomofauna, permitiendo obtener una mayor muestra para el análisis de correlación con la temperatura. No obstante, a pesar de ello, se cumplieron los objetivos del estudio, ya que se hallaron correlaciones estadísticas entre factores bióticos y abióticos, así como interacciones biológicas entre insectos.
Entomofauna asociada al cultivo
El análisis de la entomofauna asociada al cultivo reveló que el orden Hymenóptera fue el grupo predominante en todas las fases fenológicas, con un total de 109 individuos de Digonogastra sp. En menor proporción se registraron Hemiptera (57 individuos, entre Lygaeidae, Zelus spp., Sibovia spp. y Empoasca kraemeri), Diptera (39 individuos, incluyendo Toxomerus spp., O. dimidiatus y Allograpta spp.) y Coleóptera (30 individuos, correspondientes a H. convergens y C. sanguinea). Los órdenes Lepidóptera (18 individuos de Rupela albinella) y Neuróptera (6 individuos de C. externa) mostraron una representación menor (Figura 5). Al respecto, es probable que la composición de esta entomofauna haya sido influenciada por la aplicación de azufre (PD), utilizado como acaricida y fungicida para controlar ácaros y hongos como el oídio. De acuerdo con Griffith et al. (2015), el azufre es un insumo que se ha utilizado ampliamente durante siglos como compuesto básico para el control de plagas y enfermedades en la agricultura. Según Liang et al. (2025), el azufre se emplea ampliamente en fungicidas para el control de enfermedades en sistemas agrícolas. No obstante, puede tener efectos sobre artrópodos no objetivo, incluyendo insectos benéficos, lo cual se ha demostrado en insectos coccinélidos (mariquitas) (Gesraha & Ebeid, 2019). En ese sentido, los resultados obtenidos fueron relevantes y permitieron identificar diversas interacciones biológicas entre las especies registradas.
Los resultados obtenidos evidencian que la fenología del frijol caupí influye de manera decisiva en la composición y abundancia de insectos asociados, ya que en la fase vegetativa se registró la mayor riqueza general, destacando Hymenóptera y Hemíptera como los grupos más representativos. Durante la floración, la persistencia de Hymenóptera y el incremento de Díptera sugieren una mayor interacción con los recursos florales y posibles procesos de polinización. En la fase reproductiva, Hymenóptera vuelve a predominar, aunque la abundancia de otros órdenes disminuye de forma considerable, lo cual refleja un patrón en el que este grupo se consolida como constante a lo largo del ciclo, mientras que los demás insectos responden a los cambios fenológicos. Este comportamiento coincide con lo reportado en Colombia para Solanum sessiliflorum, donde la entomofauna aumentó significativamente en la floración, confirmando que la dinámica poblacional de insectos se encuentra estrechamente ligada a la fenología del cultivo (Torres et al., 2021). Esto es respaldado por Batyrshina et al. (2020), quienes demostraron qué, la maduración temprana de las plantas incrementa la presencia de pulgones, reforzando la idea de que las etapas fenológicas condicionan la vulnerabilidad frente a plagas específicas. No obstante, Gonzáles et al. (2016) y Gonzales et al. (2017), describen qué, la composición y dinámica de las comunidades de artrópodos están determinadas por la interacción entre la fenología del cultivo, la cobertura forestal del paisaje es más influyente que la proximidad a un bosque, el cual tanto la abundancia como el movimiento de insectos en la interfaz bosque–cultivo, algo visto en los cultivos de soja.
Por otro lado, Greiser et al. (2022) y Dunn et al. (2023) destacan que la variación microclimática, también tiene un rol en la modulación de la fenología de los cultivos y la sincronía con los insectos, generando respuestas diferenciales según la región. Por tanto, estos antecedentes y los resultados obtenidos en el presente estudio confirman que la fenología constituye un factor central en la estructuración de comunidades entomológicas, determinando tanto la abundancia como la función ecológica de los grupos de insectos a lo largo del ciclo del cultivo, por lo que los resultado obtenidos pueden ser dilucidados, resaltando a su vez que este estudio sería el primero que se ha realizado en cultivos de frijol caupi (V. unguiculata L.) en el norte de Perú.
CONCLUSIONES
El presente estudio evidencia que el cultivo de V. unguiculata L. bajo manejo orgánico promueve una elevada diversidad de interacciones biológicas, en las que conviven insectos polinizadores, depre-dadores, plagas, migratorios y/o visitantes y parasitoides. Los resultados demuestran que la fenología del cultivo constituye un factor deter-minante en la variación y abundancia de las especies registradas, superando en influencia al efecto de la temperatura ambiental. Estos hallazgos destacan la relevancia de considerar las etapas fenológicas para optimizar las estrategias de manejo agroecológico, minimizando el impacto de plagas y favoreciendo la conservación de insectos benéficos en sistemas agrícolas sostenibles.
Se sugiere desarrollar estudios longitudinales que abarquen múltiples ciclos de cultivo y diferentes condiciones agroecológicas, con el fin de evaluar la estabilidad temporal de las interacciones entre la entomofauna, la fenología y los factores ambien-tales. Asimismo, sería pertinente incorporar análisis moleculares y de redes ecológicas que permitan profundizar en las relaciones tróficas y funcionales entre las especies registradas.
AGRADECIMIENTOS
A la Estación Experimental Agraria (EEA) Los Cedros del Instituto Nacional de Innovación Agraria (INIA) por el apoyo brindado en la ejecución del presente estudio de investigación. A la Ing. MSc. Patricia Cruz Córdova, jefa del museo entomológico de la Facultad de Ciencias Agrarias de la Universidad Nacional de Tumbes, por su apoyo brindado en el montaje de los insectos.
REFERENCIAS BIBLIOGRÁFICAS
Abebe, B., & Alemayehu, M. (2022). A review of the nutritional use of cowpea (Vigna unguiculata L. Walp) for human and animal diets. Journal of Agriculture and Food Research, 10,100383 https://doi.org/10.1016/j.jafr.2022.100383
Álamo-Herrera, C. R., Arteaga, M. C., & Bello-Bedoy, R. (2024). Diversidad genética y variación fenotípica en una avispa parasitoide involucrada en la interacción yuca-polilla de la yuca. Revista Mexicana de Biodiversidad, 95, e955461-e955461. https://doi.org/10.22201/ib.20078706e.2024.95.5461
Alfonso-Sosa, M. I., Ardila-Bayona, V., Perez-Posada, J. J., & Rodríguez, J. S. M. (2024). Moscas de las flores (Diptera: Syrphidae) asociadas a cuatro coberturas vegetales en Ubaque, Cundinamarca. Revista Novedades Colombianas, 19(2),46-57. https://doi.org/10.47374/novcol.2024.v19.2484
Andrade, K. A., Aguiar-Menezes, E. L., Gonçalves-Esteves, V., Mendonça, C. B. F., Vieira, G. R. M., Melo, S. J., ... & Melo, G. J. B. (2017). Pollen ingestion by Chrysoperla externa (Hagen) adults in a diversified organic agroecosystem. Neotropical Entomology, 47(1), 118-130. https://doi.org/10.1007/s13744-017-0537-8
Ansar, A., Santhoshkumar, T., Radhakrishnan, G., & Jayapal, A. (2024). Impact of key insect pests on cowpea and their relationship with weather parameters. International Journal of Environment and Climate Change, 14(11), 683-692. https://doi.org/10.9734/ijecc/2024/v14i114578
Arcaya, E., Pérez-Bañón, C., Mengual, X., Zubcoff-Vallejo, J. J., & Rojo, S. (2017). Life table and predation rates of the syrphid fly Allograpta exotica, a control agent of the cowpea aphid Aphis craccivora. Biological Control, 115, 74-84. http://dx.doi.org/10.1016/j.biocontrol.2017.09.009
Baltazar, H. (2016). Factores climáticos que influyen en la diversidad de insectos en Spartium junceum L. (Fabales: Fabaceae). Prospectiva Universitaria en Ingeniería y Tecnología, 13(1), 30-48. https://doi.org/10.17843/rpmesp.2024.413.13838
Bastidas, J. S., Devia, E. H. V., & Amaya, Óscar S. (2010). Cría y evaluación de la capacidad de depredación de Chrysoperla externa sobre Neohydatothrips signifer, trips plaga del cultivo de maracuyá. Ciencia Y Tecnología Agropecuaria, 11(1), 31–40. https://doi.org/10.21930/rcta.vol11_num1_art:192
Batyrshina, Z. S., Cna’ani, A., Rozenberg, T., Seifan, M., & Tzin, V. (2020). The combined impacts of wheat spatial position and phenology on cereal aphid abundance. PeerJ, 8, e9142. https://doi.org/10.7717/peerj.9142
Cabrera-Córdoba, S. P., Gutiérrez-Torres, J. D., & Restrepo-Manrique, R. (2021). Effects of environmental variables on the composition of ants (Hymenoptera: Formicidae) in Amazonian foothills forest. Iteckne, 18(1), 57-70. https://doi.org/10.15332/iteckne.v18i1.2541
Carter, N., Powell, W., Wright, A. F., & Ashby, J. E. (1989). Effectiveness of different insecticides applied at various growth stages to control aphids on winter wheat. Crop Protection, 8(4), 271-276. https://doi.org/10.1016/0261-2194(89)90012-4
Castillo-Carrillo, P. S. (2013). Manual de plagas del cultivo de fríjol caupí. Universidad Nacional de Tumbes. https://doi.org/10.13140/RG.2.1.3971.3121
Cedillo Mozombite, K., Castillo-Carrillo, P. S., & Núñez Sacarias, E. (2020). Especies de crisópidos en plantaciones de limón sutil (Citrus aurantifolia Swingle) en la Región de Tumbes, Perú. Revista Peruana De Entomologia, 53(1 & 2), 1–10.
Coloma, D. A. C., Iznaga, Á. L. S., Gavilanez, J. R. M., Chang, G. M. C., Gaibor, P. P. R., & Marìn, E. X. C. (2023). Estudio teórico de alternativas para el control de Rupella albinella en cultivares de arroz (Oryza sativa). INQUIDE-Ingeniería Química y Desarrollo, 5(02), 01-09. https://doi.org/10.53591/iqd.v5i02.1040
Da Costa, C., Da Silva, M., De Lima Marques, A., Moura, D., Linhares, A., Oliveira, H., Moura, T., Da Silva, N., Da Silva Pontes, N., & Barbosa, M. (2024). Organic fertilization as a sustainable alternative in the production of bean varieties in the Brazilian semiarid region. Observatório de la Economía Latinoamericana, 22(2),1-21. https://doi.org/10.55905/oelv22n2-195
de Freitas Bueno, A., Batistela, M. J., de Freitas Bueno, R. C. O., de Barros França-Neto, J., Nishikawa, M. A. N., & Libério Filho, A. (2011). Effects of integrated pest management, biological control and prophylactic use of insecticides on the management and sustainability of soybean. Crop Protection, 30, 937-945. https://doi.org/10.1016/j.cropro.2011.02.021
Dewi, V., Nurariaty, A., Tuwo, M., et al. (2020). Arthropoda diversity in organic cocoa farming in Bantaeng district. In IOP Conference Series: Earth and Environmental Science, 486, 012164. https://doi.org/10.1088/1755-1315/486/1/012164
Díaz-Lucas, M. F., Maza, N., Kirschbaum, D. S., Rocca, M., & Greco, N. M. (2024). The potential of the hoverfly Allograpta exotica as a biological control agent of the strawberry aphid Chaetosiphon fragaefolii. Journal of Applied Entomology, 148(2), 191-198. https://doi.org/10.1111/jen.13220
Díaz Avalos, A. P., Leiva Chimbor, L. A., Rodríguez Rodríguez, R., Hurtado Mendoza, C. A., Rodríguez Soto, J. C., Díaz Pretell, L. E. (2021). Determinación por morfometría geométrica de Varroa sp. (Acari: Varroidae) ectoparásito de Apis mellifera L., costa norte del Perú, 2018. Arnaldoa, 28(3),717-726. http://dx.doi.org/10.22497/arnaldoa.283.28314
Díaz, J. R. H., de Barrera, L. D. C. B., Teixeira, A. A. G., Fucos, M. C. A., van Heurck, M., de Del Águila, E. R., & Flores, H. A. D. (2025). Insectos al Acecho: Dinámica Estacional en el Cultivo de Chiclayo Verdura en la Amazonía. Tecno Humanismo, 5(1), 1-77. https://doi.org/10.53673/th.v5i3.423
Doria Bolaños, M., Ramirez Navarro, M., & Fachin Ruiz, G. (2021). Ciclo biológico de Caligo idomeneus (Nymphalidae) bajo condiciones controladas con jardines verticales en Tarapoto. Revista Agrotecnológica Amazónica, 1(1), 20-33. https://doi.org/10.51252/raa.v1i1.105
Doyle, T., Hawkes, W. L., Massy, R., Powney, G. D., Menz, M. H., & Wotton, K. R. (2020). Pollination by hoverflies in the Anthropocene. Proceedings of the Royal Society B, 287(1927), 20200508. https://doi.org/10.1098/rspb.2020.0508
Dunn, P. O., Ahmed, I., Armstrong, E., Barlow, N., Barnard, M. A., Bélisle, M., & Whittingham, L. A. (2023). Extensive regional variation in the phenology of insects and their response to temperature across North America. Ecology, 104(5), e4036. https://doi.org/10.1002/ecy.4036
Ferrufino, A. (2010). Plagas del frijol. Guía de identificación y manejo integrado. https://dicta.gob.hn/files/2010,-Identificacion-y-manejo-de-plagas-del-frijol-en-CA.pdf
Funichello, M., costa, L. L., aguirre, G., & busoli, A. C. (2012). Biological aspects of Cycloneda sanguínea (Coleóptera: Coccinellidae) feeding on aphids reared on transgenic cotton Bollgard I. Revista Colombiana de Entomología, 38(1), 156–161. https://doi.org/10.25100/socolen.v38i1.8981
Gamboa, S., Souza, B., & Morales, R. (2016). Predatory activity of Chrysoperla externa (Neuroptera: Chrysopidae) on Macrosiphum euphorbiae (Hemiptera: Aphididae) in Rosa sp. crop. Revista Colombiana de Entomología, 42(1), 54-58. https://doi.org/10.25100/socolen.v42i1.6670
Gesraha, M. A., & Ebeid, A. R. (2019). Impact of sulfur dust application on the abundance of two important coccinellid predators in marrow fields. Bulletin of the National Research Centre, 43(1), 34. https://doi.org/10.1186/s42269-019-0060-7
Giorgi, J. A., Vandenberg, N. J., McHugh, J. V., Forrester, J. A., Ślipiński, S. A., Miller, K. B., Shapiro, L. R., & Whiting, M. F. (2009). The evolution of food preferences in Coccinellidae. Biological Control, 51, 215-231. https://doi.org/10.1016/j.biocontrol.2009.05.019
Godfray, H. C. J., & Chan, M. S. (1990). How insecticides trigger single stage outbreaks in tropical pests. Functional Ecology, 4(3), 329-337. https://doi.org/10.2307/2389594
Gonçalves, A., Goufo, P., Barros, A., Domínguez‐Perles, R., Trindade, H., Rosa, E., Ferreira, L., & Rodrigues, M. (2016). Cowpea (Vigna unguiculata L. Walp), a renewed multipurpose crop for a more sustainable agri-food system: Nutritional advantages and constraints. Journal of the Science of Food and Agriculture, 96(9), 2941-2951. https://doi.org/10.1002/jsfa.7644
González, E., Salvo, A., Defagó, M. T., & Valladares, G. (2016). A moveable feast: Insects moving at the forest-crop interface are affected by crop phenology and the amount of forest in the landscape. PLoS ONE, 11(7), e0158836. https://doi.org/10.1371/journal.pone.0158836
Gonzalez, E., Salvo, A., & Valladares, G. (2017). Arthropod communities and biological control in soybean fields: Forest cover at landscape scale is more influential than forest proximity. Agriculture, ecosystems & environment, 239, 359-367. https://doi.org/10.1016/j.agee.2017.02.002
Grassberger, M., & Reiter, C. (2001). Effect of temperature on Lucilia sericata (Diptera: Calliphoridae) development with special reference to the isomegalen-and isomorphen-diagram. Forensic science international, 120(1-2), 32-36. https://doi.org/10.1016/S0379-0738(01)00413-3
Greiser, C., von Schmalensee, L., Lindestad, O., Gotthard, K., & Lehmann, P. (2022). Microclimatic variation affects developmental phenology, synchrony and voltinism in an insect population. Functional Ecology, 36(12), 3036-3048. https://doi.org/10.1111/1365-2435.14195
Griffith, C. M., Woodrow, J. E., & Seiber, J. N. (2015). Environmental behavior and analysis of agricultural sulfur. Pest management science, 71(11), 1486-1496. https://doi.org/10.1002/ps.4067
Horgan, FG (2023). La estructura de las comunidades de barrenadores del tallo del arroz: una revisión de las distribuciones de especies, rangos de hospedadores e interacciones interespecíficas. Insects, 14(12), 921. mhttps://doi.org/10.3390/insects14120921
Hopper, J. V., Nelson, E. H., Daane, K. M., & Mills, N. J. (2011). Growth, development and consumption by four syrphid species associated with the lettuce aphid, Nasonovia ribisnigri, in California. Biological Control, 58(3), 271-276. https://doi.org/10.1016/j.biocontrol.2011.03.017
Jackai, L. E. N., & Daoust, R. A. (1986). Insect pests of cowpeas. Annual Review of Entomology, 31, 95-119. https://doi.org/10.1146/annurev.en.31.010186.000523
Jankielsohn, A. (2018). The importance of insects in agricultural ecosystems. Agricultural Ecosystems, 06, 62-73. https://doi.org/10.4236/ae.2018.62006
Jasrotia, P., Kumari, P., Malik, K., Kashyap, P., Kumar, S., Bhardwaj, A., & Singh, G. (2023). Conservation agriculture-based crop management practices impact diversity and population dynamics of the insect-pests and their natural enemies in agroecosystems. Frontiers in Sustainable Food Systems, 7, 1173048. https://doi.org/10.3389/fsufs.2023.1173048
Khalifa, S.A.M., Elshafiey, E.H., Shetaia, A.A., El-Wahed, A.A.A., Algethami, A.F., Musharraf, S.G., Alajmi, M.F., Zhao, C., Masry, S.H.D., Abdel-Daim, M.M., Halabi, M. F., Kai, G., Al Naggar, Y., Bishr, M., Diab, M.A.M., El-Seedi, H.R., (2021). Overview of bee pollination and its economic value for crop production. Insects, 12(8). https:// doi.org/10.3390/insects12080688.
Lazaridi, E., Suso, M. J., Ortiz-Sánchez, F. J., & Bebeli, P. J. (2023). Investigation of cowpea (Vigna unguiculata (L.) Walp.)–insect pollinator interactions aiming to increase cowpea yield and define new breeding tools. Ecologies, 4(1), 124-140. https://doi.org/10.3390/ecologies4010010
Lehmann, P., Ammunét, T., Barton, M., Battisti, A., Eigenbrode, S. D., Jepsen, J. U., & Björkman, C. (2020). Complex responses of global insect pests to climate warming. Frontiers in Ecology and the Environment, 18(3), 141-150. https://doi.org/10.1002/fee.2160
Leite, G. L. D., Silva, F. W. S., Ferreira, C. S., Guañabens, R. E. M., Lazo, J. A., Sampaio, R. A., & Fonseca, I. M. (2011). Intensidad de plagas y sus enemigos naturales en una asociación de maíz y fríjol fertilizados con diferentes formas de lodo de alcantarillado. Arquivos do Instituto Biológico, 78(3), 367-375. https://doi.org/10.1590/1808-1657v78p3672011
Liang, X., Zhang, Y., Wan, S., Xia, L., Li, P., Wang, S., ... & Zhang, Y. (2025). Impacts of sulfur application on microbial communities and functional attributes in rubber plantation soil. BMC microbiology, 25(1), 265. https://doi.org/10.1186/s12866-025-03971-z
Liccardo, A., Fierro, A., Garganese, F., Picciotti, U., & Porcelli, F. (2020). A biological control model to manage the vector and the infection of Xylella fastidiosa on olive trees. PLoS One, 15, e0232363. https://doi.org/10.1371/journal.pone.0232363
Lima, L. R., de Sá Cavalcanti, W. H., Junior, C. M. J. B., & Silva, R. C. (2022). Modificação mecânica de um aspirador de pó convencional em aspirador entomológico para Isoptera. Revista Eletrônica Perspectivas da Ciência e Tecnologia, 14, 1-22. https://doi.org/10.22407/1984-5693.2022.v14.p.e20221404
Lobato Rodrigues, N. E., da Silva, A. G., de Souza, B. H. S., Costa, E. N., Ribeiro, Z. A. & Boiça Júnior, A. L. (2015). Effects of cowpea cultivars and neem oil on attractiveness, feeding, and development of Spodoptera eridania (Cramer) (Lepidoptera: Noctuidae). IDESIA, 33(4), 65-74. http://dx.doi.org/10.4067/S0718-34292015000400009
Lucas, M. F., Passareli, L. M., Maza, N., Aquino, D. A., Greco, N. M., & Rocca, M. (2020). Variación espacio-temporal de sírfidos depredadores (Díptera: Syrphidae) y su asociación con áfidos en cultivos hortícolas orgánicos de La Plata, Buenos Aires. Revista de la Sociedad Entomológica Argentina, 79(4), 15-22. https://doi.org/https://doi.org/10.25085/rsea.790403.
Macchia, G. (2024). Control biológico por conservación: conceptos clave, estrategias y avances para construir una agroecología sostenible. Revista de la Facultad de Agronomía, 123(1), 143-143. https://doi.org/10.24215/16699513e143
Mateos‐Fierro, Z., Leach, A., & Kaplan, I. (2025). Insecticides outweigh rye cover crop in triggering secondary pest outbreaks. Pest Management Science, 81(12), 7971-7984. https://doi.org/10.1002/ps.70109
Metcalf, R. L. (1980). Changing role of insecticides in crop protection. Annual Review of Entomology, 25(1), 219-256. https://doi.org/10.1146/annurev.en.25.010180.001251
Mochizuki, M. (2003). Effectiveness and pesticide susceptibility of the pyrethroid-resistant predatory mite Amblyseius womersleyi in the integrated pest management of tea pests. BioControl, 48(2), 207-221. https://doi.org/10.1023/A:1022673317471
Mourou, M., Incampo, G., Carlucci, M., Salamone, D., Pollastro, S., Faretra, F., & Nigro, F. (2025). Insight into biological strategies and main challenges to control the phytopathogenic bacterium Xylella fastidiosa. Frontiers in Plant Science, 16, 1608687. https://doi.org/10.3389/fpls.2025.1608687
Moya-Raygoza, G. (2005). Relationships between the ant Brachymyrmex obscurior (Hymenoptera: Formicidae) and Acacia pennatula (Fabaceae). Insectes Sociaux, 52(2), 105-107. https://doi.org/10.1007/s00040-004-0777-6
Murgas, A. S. (2024). Aspectos biológicos de Ocyptamus gastrostactus y controlador biológico Pachyneuron albutius, en Panamá. The Biologist, 22(1), 85-94. https://doi.org/10.62430/rtb20242211684
Nadal, M. F., Avalos, G., & González, A. (2024). Diversidad de arañas saltadoras (Araneae: Salticidae) en hábitats del Chaco Oriental de Argentina con diferentes estados de conservación. Revista Chilena de Entomología, 50(2), 259-308. https://doi.org/10.35249/rche.50.2.24.13
Naranjo-Acosta, J., Díaz-Mesa, D. C., Melo-Cerón, C. I., & Manzano, M. R. (2022). Depredación, reproducción y desarrollo de la catarina Hippodamia convergens (Coleoptera: Coccinellidae). Revista de Biología Tropical, 70(1), 621-635. http://dx.doi.org/10.15517/rev.biol.trop.2022.48371
Nehybová, K., & Horak, J. (2024). Biodiversity of pollinators in agricultural landscapes and the role of weather and conservation management. Biological Conservation, 300, 110875. https://doi.org/10.1016/j.biocon.2024.110875
Outhwaite, C. L., McCann, P., & Newbold, T. (2022). Agriculture and climate change are reshaping insect biodiversity worldwide. Nature, 605(7908), 97-102. https://doi.org/10.1038/s41586-022-04644-x
Plataforma del Estado Peruano. (s.f.). Tumbes. http://met.igp.gob.pe/clima/HTML/tumbes.html
Ramos Martínez, J. A., Bautista Martínez, N., Guzmán Franco, A. W., Rodríguez Maciel, J. C., Trujillo Arriaga, F. J., & Tamayo Mejía, F. (2023). Morphological and Molecular Identification of Diatraea grandiosella Dyar and its Natural Enemies Associated with Maize Cultivation at Guanajuato, Mexico. Southwestern Entomologist, 48(49, 925-938. https://doi.org/10.3958/059.048.0415
Reguilón, C., Olivares, N., & Pérez, Y. (2023). Primer reporte de dos especies de Chrysoperla Steinmann, 1964 (Neuroptera: Chrysopidae) asociadas a cítricos en Chile. Revista Chilena de Entomología, 49(4), 735-739. http://dx.doi.org/10.35249/rche.49.4.23.07
Rojas-Pérez, F., López, D. J. P., Obrador-Olán, J. J., Salgado-García, S., Palma-Cancino, D. J., Arreola-Enríquez, J., & Huicab-Pech, Z. G. (2025). Rendimiento y calidad nutricional de frijol castilla (Vigna unguiculata L. Walp) bajo la aplicación de fertilizantes orgánicos líquidos. Agro Productividad, 3,192-202. https://doi.org/10.32854/b9wvg842
Ruiz-Polo, A. A., & Barrera-Rivera, L. V. (2024). Efecto de la temperatura en el ciclo biológico del Aedes aegypti en condiciones de laboratorio. Revista Peruana de Medicina Experimental y Salud Pública, 41(3), 327-328. http://dx.doi.org/10.17843/rpmesp.2024.413.13838
Ruiz-Sánchez, E., Ruiz-Jiménez, A. L., Latournerie-Moreno, L., & Pierre, J. F. (2025). Insecticidas químicos y biorracionales utilizados por pequeños productores de hortalizas de Yucatán. Avances en Investigación Agropecuaria, 29, 130. https://doi.org/10.53897/RevAIA.25.29.48
Saleem, M., Fatima, A., Rehman, A., Aroob, T., Aleem, A., Shahbaz, M., Naveed, H., Majeed, A., Nawaz, M., & Gulsher, M. (2024). Insect symphony: Understanding the role of entomology in ecosystem harmony. Journal of Physics and Biomedical Biological Science, 2024(1), 26-26. https://doi.org/10.64013/jpbab.v2024i1.26
Salas-Marina, M. A., Hernández-García, V., Ordaz-Silva, S., Delgadillo-Ángeles, J. L., López-Sánchez, I. V., Chacón-Hernández, J. C., & Soto-Hernández, M. (2023). Natural parasitism of Monoctenus sanchezi (Hymenoptera: Diprionidae: Monocteninae) in the Sierra de Álvarez, San Luis Potosí, México. Southwestern Entomologist, 47(4), 829-834. https://doi.org/10.3958/059.047.0404
Sanchez-Alvarado, E., Herrera-Reyes, S., Arellano, C. S., Luna, F. G., Valarezo-Rivera, N., & Valencia, P. E. (2023). Monitoreo de insectos plaga mediante SIG aplicados al cultivo de Oryza sativa L. en Naranjal, Ecuador. Manglar, 20(1), 59-67. http://dx.doi.org/10.57188/manglar.2023.007
Sahani, S., Chandra, K., Girase, I., Saikanth, D., Singh, S., Swamy, G., & Singh, B. (2023). Role of insect pollinators in crop production and ecosystem. Uttar Pradesh Journal of Zoology, 44(18),42-52. https://doi.org/10.56557/upjoz/2023/v44i183604
Sharma, T., Singh, J., Madaik, S., Kumar, P., Singh, A., Rana, B., & Chauhan, G. (2024). Organic input incorporation for enhancing sustainability and economic viability of cowpea in North-Western Himalayan region. Frontiers in Agronomy, 6,1458603. https://doi.org/10.3389/fagro.2024.1458603
Santos-Murgas, A., Toribio, E. J., & Cobos-Hernández, R. M. (2024). Biología de Syntomeida melanthus (Lepidoptera: Erebidae) en el sitio Ramsar, Humedal Bahía de Panamá. I+D Tecnológico, 20(1), 5-9. https://doi.org/10.33412/idt.v20.1.3833
Schneider, L., Rebetez, M., & Rasmann, S. (2022). The effect of climate change on invasive crop pests across biomes. Current Opinion in Insect Science, 50, 100895. https://doi.org/10.1016/j.cois.2022.100895
Servicio Nacional de Meteorología e Hidrología del Perú (SENAMHI). (2025). Pronóstico del tiempo para Tumbes. https://www.senamhi.gob.pe
Skendžić, S., Zovko, M., Živković, I. P., Lešić, V., & Lemić, D. (2021). The impact of climate change on agricultural insect pests. Insects, 12(5), 440. https://doi.org/10.3390/insects12050440
Sturza, V. S., Dequech, S. T. B., Toebe, M., Silveira, T. R., Cargnelutti Filho, A., & Bolzan, A. (2014). Toxomerus duplicatus Wiedemann, 1830 (Diptera: Syrphidae) preying on Microtheca spp. (Coleoptera: Chrysomelidae) larvae. Brazilian Journal of Biology, 74(3), 656-658. https://doi.org/10.1590/bjb.2014.0071
Subedi, B., Poudel, A., & Aryal, S. (2023). The impact of climate change on insect pest biology and ecology: Implications for pest management strategies, crop production, and food security. Journal of Agriculture and Food Research, 14, 100733. https://doi.org/10.1016/j.jafr.2023.100733
Torres-Torres, J. J., Murillo-Barahona, D. A., & Rengifo-Murillo, L. (2021). Entomofauna asociada al cultivo de Solanum sessiliflorum Dunal, en el municipio de Lloró, occidente de Colombia. Revista Politécnica, 17(33), 90-99. https://doi.org/10.33571/rpolitec.v17n33a7
Urías-Rivas, M. O., Benitez-Dieguez, I., & Bojórquez-López, M. (2024). Efectos de las prácticas de agroquímicos en la salud humana y el medio ambiente en la agricultura sostenible. Revista Ra Ximhai, 20(2), 151-171. https://doi.org/10.35197/rx.20.02.2024.07.mu
Valarezo Beltrón, C. O., Saldarriaga Lucas, V. A., Vera Montenegro, L., Julca Otiniano, A., & Rodríguez Berrío, A. (2022). Diversidad de insectos en el cultivo de Citrus aurantifolia y vegetación aledaña en dos localidades ecológicas de Ecuador. Revista de Investigación e Innovación Agropecuaria y de Recursos Naturales, 9(2), 44-51. https://doi.org/10.53287/cgcj2651ps25z
Valente, E. C. N., Broglio, S. M. F., Dias-Pini, N. da S., Lima, A. S. T. de, & Micheletti, L. B. (2018). (Record of Digonogastra sp. (Hymenoptera: BRACONIDAE) parasitizing Diatraea flavipennella (Lepidoptera: Crambidae) in Alagoas, Brazil. Revista Caatinga, 31(2), 511–514. http://dx.doi.org/10.1590/1983-21252018v31n228rc
Vaidya, C., Cruz, M., Kuesel, R., Gonthier, D. J., Iverson, A., Ennis, K. K., & Perfecto, I. (2017). Local and landscape constraints on coffee leafhopper (Hemiptera: Cicadellidae) diversity. Journal of Insect Science, 17(2), 38. https://doi.org/10.1093/jisesa/iew127
Vidal, M. C., Abarca, M., Backe, K., Curé, A. E., Finke, D. L., Koltz, A. M., & Murphy, S. M. (2025). ¿Qué sabemos sobre las respuestas de los insectos al cambio global? Una revisión de metaanálisis sobre los factores que impulsan el cambio global. Conservación y Diversidad de Insectos, 10(3), 12845. https://doi.org/10.29166/siembra.v9i3
Vivas-Carmona, L. E., & Astudillo García, D. H. (2022). Fluctuación poblacional de la novia del arroz (Rupela albinella) en Calabozo, Estado Guárico, Venezuela. Journal of the Selva Andina Biosphere, 10(1), 7-20. https://doi.org/10.36610/j.jsab.2022.100100007