 |
INTRODUCCIÓN
Dinámica del ciclo celular y determinación de la hora mitótica en tres razas peruanas de maíz a partir de meristemos radiculares
Cell cycle dynamics and determination of the mitotic hour in three Peruvian maize races from root meristems
Valeryn Cruz1; Lady Trinidad1; María Siles1; Alberto López1*
1 Laboratorio de Genética. Grupo de Investigación en Recursos Genéticos (RECGEN), Facultad de Ciencias Biológicas, Universidad Nacional Mayor de San Marcos. Lima 15081, Lima, Perú.
* Autor corresponsal: alopezs@unmsm.edu.pe (A. López).
ORCID de los autores
V. Cruz: https://orcid.org/0009-0006-2678-7800 L. Trinidad: https://orcid.org/0009-0004-5316-4663
M. Siles: https://orcid.org/0000-0003-4956-8310 A. López: https://orcid.org/0000-0001-6070-5836
RESUMEN
Palabras clave: Zea mays, razas peruanas; hora mitótica; ciclo celular; ciclo mitótico.
ABSTRACT
Maize (Zea mays L.) is one of the most widely cultivated cereals worldwide, with great agronomic, industrial, nutritional, and cultural importance. It is also considered a classical model in cytogenetic studies due to its genetic diversity and chromosomal structure. In such analyses, determining the cell cycle dynamics and mitotic hour is crucial for obtaining high-quality chromosome preparations; however, this aspect remains insufficiently explored in maize. In this context, a cytogenetic slide preparation protocol was optimized to assess cell cycle dynamics and determine the mitotic hour in three Peruvian maize races: Confite puntiagudo, Piricinco, and Cuzco gigante var. Blanco Urubamba. Clean and uniformly stained slides were obtained using the acid–enzymatic hydrolysis treatment. The mitotic hour was recorded around 12:00 h in Confite puntiagudo and Blanco Urubamba, whereas no precise range was defined for Piricinco. The duration of the mitotic cycle was similar among races (0.42 – 0.49 h), suggesting a conserved cell cycle dynamic.
Keywords: Zea mays; Peruvian races; mitotic hour; cell cycle; mitotic cycle.
Recibido: 21-12-2025.
Aceptado: 21-03-2026.
El maíz (Zea mays L.), perteneciente a la familia Poaceae, es junto al trigo y al arroz uno de los cereales más cultivados a nivel mundial (García Mendoza, 2017). Se distingue por su alto potencial genético de rendimiento, buen valor económico y una amplia adaptabilidad a diversos climas y tipos de suelos, características que han favorecido su expansión global (Sanodiya et al., 2023).
Además de su relevancia agrícola e industrial, desempeña un papel esencial en la seguridad alimentaria, la economía global y en la cultura y tradición de diversos países de África y América Latina (Palacios-Rojas et al., 2020; Song et al., 2025; Kim et al., 2026). Es considerado también como un alimento de alta calidad para animales, sobre todo rumiantes (Torres et al., 2025). Su domesticación data de hace aproximadamente 9000 años en el suroeste de México, a partir de su pariente silvestre, el teocinte (Liu et al., 2020; Panda et al., 2020; Domínguez et al., 2024). Posteriormente, se dispersó hacia la región andina de Sudamérica donde se han reconocido 146 razas (Salvador-Reyes & Clerici, 2020), siendo el Perú considerado como un centro de diversificación del cultivo (MINAM, 2018), identificándose alrededor de 52 razas de maíz (Fuentes-Cárdenas et al., 2022) clasificadas principalmente según sus rasgos morfológicos, ubicación geográfica y proceso evolutivo (Salhuana, 2003). Arias et al. (2025) refieren que la conservación y protección de estas razas nativas deben priorizarse, promoviéndose mayor cantidad de estudios sobre bioseguridad, conservación de su biodiversidad entre otros.
Asimismo, el maíz presenta una gran diversidad genética, relacionada con factores geográficos, ecológicos y culturales, que han permitido la obtención de genotipos mejorados (Barrera-Guzmán et al., 2020; Cheng & Wang, 2025). Por esta razón, es considerado un organismo modelo para la investigación básica, principalmente en las áreas de citología, genética, genómica y mejoramiento vegetal, debido a su desarrollo floral monoico y heterólogo, sincronía meiótica y tamaño grande y distintivo de sus cromosomas (Hake & Ross-Ibarra, 2015). Zambrano et al. (2025) plantean que la investigación y conservación de la semilla nativa del maíz debe promoverse, dado que es una fuente de diversidad genética y es donde se encontrarían los genes que pueden hacer frente al cambio climático.
Entre los estudios citogenéticos realizados en maíz destacan aquellos orientados a la determinación del número cromosómico, nivel de ploidía, elaboración de cariotipos, evaluación de genotoxicidad y mapeo génico. Un paso previo fundamental en estos análisis es la selección y optimización del método de preparación de láminas citogenéticas, ya que de ello depende una adecuada visualización de las células al microscopio.
Diversos autores han descrito protocolos de preparación de láminas citogenéticas empleando diferentes fijadores, agentes de hidrólisis y colorantes (McCaw et al., 2016; Mondin et al., 2014) así como técnicas de goteo para obtener cromosomas dispersos (Yao et al., 2020).
En contraste, pocos estudios en maíz han abordado el análisis del ciclo celular y la determinación de la hora mitótica, entendida como el momento de máxima actividad de división celular, como parte esencial del proceso de estandarización, a diferencia de lo reportado en otras especies donde constituye un componente clave de la metodología para la obtención de preparados cromosómicos de alta calidad (Burbano et al., 2011; García-Godos Alcázar et al., 2020; Mata-Sucre et al., 2017; Quija-Lamina et al., 2010; Rondón et al., 2013). Kaduchová et al (2023) precisan que, en general, la división celular mitótica, su regulación, cinética y dinámica han sido estudiados con gran detalle en hongos, diversos metazoos y humanos, sin embargo, dichos estudios en plantas son aún muy limitados. Asimismo, resulta fundamental estimar los parámetros del ciclo celular en plantas para poder comprender las etapas y programas de desarrollo en la formación del organismo (Echevarría et al., 2021).
Considerando ello, el empleo de raíces como material vegetal resulta especialmente adecuado para el estudio de la proliferación celular, ya que los procesos de división y expansión celular se encuentran separados espacial y temporalmente, concentrándose la actividad mitótica en la zona meristemática (Desvoyes et al., 2021). Esta región presenta una alta proporción de células en fase M lo que evidencia una intensa proliferación celular y una dinámica de ciclo celular activa, en concordancia con análisis recientes de transcriptómica unicelular donde se caracterizó la distribución de las fases del ciclo celular en distintas subpoblaciones celulares de raíces de Zea mays (Bian et al., 2025).
Así, el estudio del ciclo celular y división mitótica en meristemos radicales de maíz es clave para comprender la dinámica y duración del ciclo celular y sus diferentes fases, aspectos que pueden variar entre genotipos debido a diferencias genéticas, fisiológicas o ambientales (Wang et al., 1986). Asimismo, conocer la hora mitótica permite definir el horario más adecuado para la recolección de las muestras (Talledo & Escobar, 1995) y mejorar la eficacia de los tratamientos químicos y físicos empleados para la obtención de cromosomas dispersos (Ferrer Pereira et al., 2007), aspecto que resulta fundamental en estudios de citogenética clásica, como la elaboración de cariotipos, y molecular, como el mapeo génico mediante hibridación fluorescente in situ (FISH) (Harun et al., 2023; Khalaf et al., 2023). Por su parte, Anissa & Widodo (2021) señalan que es importante realizar el análisis a diferentes horas de corte ya que permite explorar con más detalle el momento adecuado para obtener células en mitosis y así encontrar el tiempo efectivo de división mitótica.
No obstante, a pesar de la relevancia metodológica de la determinación de la hora mitótica y del análisis de la dinámica del ciclo celular para la optimización de protocolos citogenéticos, en maíz existe escasa información sistematizada que carac-terice estos parámetros. Los estudios específicos disponibles son limitados y en su mayoría corresponden a trabajos clásicos realizados en líneas endogámicas (Verma & Lin, 1979), sin que se hayan desarrollado análisis orientados a razas nativas o germoplasma peruano.
No existen reportes de protocolos estandarizados que determinen la hora de máxima actividad de división celular en razas específicas de maíz y su relación con la dinámica celular, generando un vacío de información que debe cubrirse teniendo en cuenta que nuestro país es centro de diversificación del maíz; ello limita la optimización de estudios citogenéticos específicos.
El objetivo del presente estudio fue estandarizar un protocolo de preparación de láminas citogenéticas para estudiar la dinámica del ciclo celular y determinar la hora mitótica de tres razas peruanas de maíz a fin de aportar información metodológica útil para futuras investigaciones en diversos campos de la citogenética vegetal.
METODOLOGÍA
Obtención del material biológico
Se emplearon semillas de tres razas peruanas de maíz (Confite puntiagudo, Piricinco y Cuzco gigante var. Blanco Urubamba), proporcionadas por el Banco de Germoplasma del Programa de Investigación y Proyección Social en Maíz de la Universidad nacional Agraria La Molina (UNALM). La selección de las razas consideró sus diferencias evolutivas, morfológicas y ecológicas (MINAM, 2018) a fin de establecer una comparación representativa para el análisis de la hora mitótica y de la dinámica del ciclo celular. Todos los procedimientos experimentales se realizaron en el Laboratorio de Genética de la Facultad de Ciencias Biológicas de la Universidad Nacional Mayor de San Marcos (UNMSM).
Germinación de las semillas
Las semillas se esterilizaron según Sauer y Burroughs (1986), con ligeras modificaciones: dos lavados en etanol al 96 % por 10 s, inmersión en NaOCl al 4 % (pH 11,2) por 1 min y enjuague en agua destilada estéril por 10 s, todos bajo agitación mediante un mezclador tipo vortex (Labnet, S0200-230V-EU, EE. UU.). Luego, se dejaron reposar 5 min y se sembraron sobre una capa de algodón húmedo dispuesta en placas Petri, a 25 °C y con un fotoperiodo de 12 h luz/12 h oscuridad.
Determinación de la hora mitótica
Se estandarizó la preparación de láminas citogenéticas a partir de los protocolos descritos por Aguilera et al. (2016) y Aliyeva-Schnorr et al. (2015). Se seleccionaron semillas germinadas con raíces de 1 – 1,5 cm de longitud (Figura 1), cuyos ápices radiculares se cortaron y recolectaron a las 10:00, 11:00, 12:00, 13:00, 14:00 y 15:00 h. Para cada hora evaluada se prepararon láminas por triplicado.
Fijación
La fijación de los ápices radiculares se realizó en etanol absoluto: ácido acético glacial (3:1) durante 24 h a −20 °C. Posteriormente, se realizaron dos lavados en agua destilada fría (5 min cada uno), seguido de dos lavados en buffer citrato 0,01 M pH 4,8 (5 min cada uno), todos bajo agitación. Posteriormente, los ápices radiculares pasaron al tratamiento de hidrolisis.
Hidrólisis
Se evaluaron tres tipos de hidrólisis: ácida, enzimática y ácido-enzimática (Tabla 1).
§ Ácida: Se utilizó HCl 1N y 5N, seguido de tres lavados en buffer citrato 0,01 M (pH 4,8; 5 min cada uno). La aplicación de la solución TARGA (acético glacial: ácido láctico: agua destilada, 9:5:6) se evaluó como tratamiento posterior a la hidrólisis con HCl, realizando tres lavados antes y después de su aplicación (5 min cada uno).
§ Enzimática: Se aplicó una solución de celulasa al 2% y pectinasa líquida al 20% (ambas de Aspergillus niger) disueltas en glicerol al 40% en buffer citrato 0,01 M (pH 4,8). Tras la digestión, se realizaron cinco enjuagues en buffer citrato (5 min cada uno).
§ Ácido-enzimática: Se combinaron HCl 5N y la solución enzimática, realizándose tres lavados en buffer citrato (5 min cada uno) tras el HCl 5N y cinco lavados en el mismo buffer tras la enzima. Cuando se invirtió el orden de aplicación, también se invirtieron los lavados correspondientes.
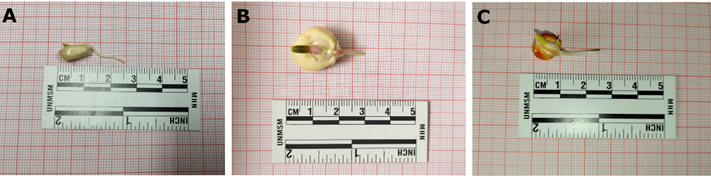
Figura 1. Semillas germinadas de tres razas peruanas de maíz, mostrando raíces de aproximadamente 1 – 1,5 cm A. Confite puntiagudo. B. Cuzco gigante var. Blanco Urubamba. C. Piricinco.
Tinción y montaje
Se evaluaron tres diferentes tiempos de tinción con orceína lactoacética al 2% (OLA 2%) —30 min, 45 min, 1 h y 1h 30 min— para cada tratamiento de hidrólisis. Transcurrido estos tiempos, se aplicó la técnica de squash y las preparaciones se sellaron con Entellan.
Conteo de células
Las láminas se observaron al microscopio óptico (Euromex iScope series, Países Bajos) a 400×, contabilizándose aproximadamente 1000 células por preparación. Las células se clasificaron de acuerdo con la fase del ciclo celular en la que se encontraban (interfase, profase, metafase, anafase o telofase).
Cálculo de índices
Se calcularon los índices de fase parciales (IF) e índice mitótico parcial (IM), de acuerdo con Talledo & Escobar (1995):
IF (%) = Número de células de cada fase / Número total de células x 100
IM (%) = IFp + IFm +IFa + IFt
Donde p: profase, m: metafase, a: anafase y t: telofase. Los índices de fase totales (IFT) se calcularon como el promedio de los IF obtenidos en cada hora de evaluación, y el índice mitótico total (IMT) como la suma de los IFT de cada fase:
IFT (%) = Suma de los IF de cada fase / Número de horas
IMT (%) = IFTp + IFTm +IFTa + IFTt
Donde p: profase, m: metafase, a: anafase y t: telofase.
La duración de cada fase del ciclo celular (TF) se estimó a partir de los IFT, asumiendo que el valor total de los IFT (100%) correspondió a la duración del ciclo celular (CC) media, equivalente al intervalo de tiempo de evaluación (5h). La duración del ciclo mitótico (CM) se obtuvo como la suma de las duraciones de las fases:
TF (h) = IFT / 100 x CC
CM (h) = TFp + TFm +TFa + TFt
Donde p: profase, m: metafase, a: anafase y t: telofase.
Análisis estadístico
El procesamiento de los datos se realizó en RStudio. Se calcularon estadísticas descriptivas (media ± desviación estándar) y se aplicó un análisis de varianza (ANOVA) de una vía. En caso de significancia (p < 0,05), se efectuó la prueba paramétrica TukeyHSD para comparaciones múltiples.
Tabla 1
Tratamientos de hidrólisis evaluados en la estandarización del protocolo de preparación de láminas citogenéticas de maíz
|
Hidrólisis |
Agente |
Temperatura |
Tiempo |
|
Ácida |
HCl 1N |
T° ambiente |
10 min |
|
20 min |
|||
|
HCl 5N |
T° ambiente |
3 min |
|
|
HCl 5N + TARGA |
T° ambiente + 4 °C |
20 min + 10 min |
|
|
20 min + 15 min |
|||
|
25 min + 15 min |
|||
|
25 min + 25 min |
|||
|
60 °C + 4 °C |
20 min + 10 min |
||
|
25 min + 10 min |
|||
|
Enzimática |
Celulasa 2% + Pectinasa 10% |
37 °C |
1h 30 min |
|
2h |
|||
|
Ácido - Enzimática |
HCl 5N + (Celulasa 2% + Pectinasa 10%) |
T° ambiente + 37 °C |
10 min + 1h 30 min |
|
15 min + 1h 30 min |
|||
|
(Celulasa 2% + Pectinasa 10%) + HCl 5N |
37 °C + T° ambiente |
1h 30 min + 10 min |
|
|
1h 30 min + 15 min |
RESULTADOS Y DISCUSIÓN
Estandarización de láminas citogenéticas
En la Tabla 2 se presenta el protocolo estanda-rizado para obtener láminas citogenéticas a partir de meristemos radiculares de maíz.
El tratamiento de hidrólisis ácida, con o sin adición de la solución ablandadora TARGA, empleada para facilitar la maceración del tejido y la separación celular, dio lugar a preparaciones con abundantes restos celulares y tinción inadecuada, lo que dificultó la distinción de células en interfase y profase. Por otro lado, el empleo exclusivo de solución enzimática mejoró ligeramente la tinción, pero produjo abundante suciedad y vesículas, sin diferencias entre los tiempos de exposición evaluados (1 h 30 min y 2 h). En contraste, la combinación de ambos tratamientos —hidrólisis enzimática por 1h 30 min seguido de hidrólisis con HCl 5N durante 15 min— produjo los mejores resultados, obteniéndose preparaciones limpias que mostraron campos con buena tinción y en las que fue posible diferenciar claramente las fases de la mitosis. Por el contrario, aplicar primero el HCl 5N y luego la enzima resultó en láminas con suciedad y presencia de vesículas.
Respecto a la tinción con OLA 2%, en las láminas sometidas únicamente a hidrólisis ácida esta resultó deficiente, sin variaciones notables entre los tiempos evaluados (30 min, 45 min, 1 h y 1 h 30 min). En aquellas tratadas solo con enzima se observó una ligera mejoría, aunque sin diferencias marcadas al prolongar el tiempo de tinción. En contraste, en las láminas donde se empleó hidrólisis combinada, la calidad de la tinción aumentó progresivamente con el tiempo de exposición al colorante, alcanzándose los mejores resultados con 1 h 30 min, donde las preparaciones se mostraron más nítidas y uniformes (Figura 2).
Tabla 2
Protocolo estandarizado de preparación de láminas citogenéticas a partir de meristemos radiculares de maíz
|
Procedimiento |
Agente |
Temperatura |
Tiempo |
Condición |
|
Fijación |
Etanol absoluto: Ácido acético (3:1) |
-20 °C |
24h |
Oscuridad |
|
Lavado |
Agua destilada |
4 °C |
2 x 5min |
Luz y agitación |
|
Buffer citrato 0,01M, pH 4,8 |
4 °C |
2 x 5 min |
||
|
Hidrólisis enzimática |
Celulasa 2% + Pectinasa 10% |
37 °C |
1h 30 min |
Luz |
|
Lavado |
Buffer citrato 0,01M, pH 4,8 |
4 °C |
5 x 5 min |
Luz y agitación |
|
Hidrólisis ácida |
HCL 5N |
Temperatura ambiente |
15 min |
Luz |
|
Lavado |
Buffer citrato 0,01M, pH 4,8 |
4 °C |
3 x 5 min |
Luz y agitación |
|
Tinción |
Orceína lactoacética 2% |
Temperatura ambiente |
1h 30 min |
Luz |
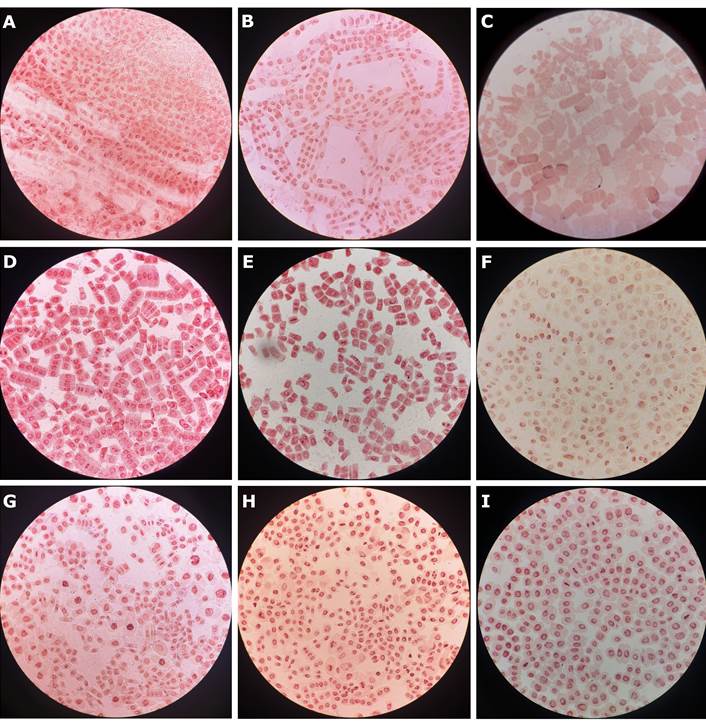
Figura 2. Células de meristemo radicular de maíz obtenidas con los tratamientos de hidrólisis evaluados durante la estandarización del protocolo de preparación de láminas citogenéticas. Microfotografías obtenidas a 400×. A. HCl 5N (3 min) B. HCl 5N (20 min, temperatura ambiente) + TARGA (15 min) C. HCl 5N (20 min, 60 °C) + TARGA (15 min) D. Hidrólisis enzimática (1 h 30 min) E. Hidrólisis enzimática (2 h) F. HCl 5N (10 min) + Hidrólisis enzimática (1 h 30 min) G. HCl 5N (15 min) + Hidrólisis enzimática (1 h 30 min) H. Hidrólisis enzimática (1 h 30 min) + HCl 5N (10 min) I. Hidrólisis enzimática (1 h 30 min) + HCl 5N (15 min).
La hidrólisis constituye un paso clave en la preparación de láminas citogenéticas, ya que permite romper la pared celular y degradar la pectina que se encuentra en las uniones intercelulares, lo que facilita la dispersión de las células en una sola capa y reduce la superposición durante la observación al microscopio (Valladolid et al., 2004). Este proceso puede realizarse mediante agentes químicos, tratamientos enzimá-ticos, o combinaciones de ambos (Valladolid et al., 2004).
Entre los agentes hidrolíticos químicos, el HCl ha sido ampliamente utilizado en diversas especies, ya sea a bajas concentraciones y temperatura ambiente (Mata-Sucre et al., 2017), o a altas temperaturas (Deogade & Nasare, 2016; Jaimes Méndez et al., 2022; Quija-Lamina et al., 2010). Asimismo, se ha reportado el empleo de HCl a mayor concentración y temperatura ambiente, con resultados satisfactorios (Burbano et al., 2011).
En el presente estudio, la hidrólisis ácida empleando HCl a diferentes concentraciones y tiempos, así como en combinación con soluciones ablandadoras como TARGA, produjo preparados con abundantes restos celulares y tinción deficiente, aun cuando se incrementó el tiempo de exposición al colorante. Esto sugiere una digestión inadecuada de la pared celular, lo que dificultó la entrada del colorante en las células.
Se ha señalado que el HCl disuelve las sales pécticas de la lámina media, facilitando la separación de las células y el aclaramiento del citoplasma (Valladolid et al., 2004), además de suavizar los ápices radiculares y favorecer el squash (Delgado et al., 2010). Sin embargo, su efecto puede ser adecuado para ciertos grupos vegetales, pero limitado para otros, particularmente especies con cromosomas pequeños y numerosos (Kawano, 1965). Lo anterior fue corroborado por Rondón et al. (2013) quienes, al emplear hidrólisis ácida en ápices radiculares de una especie de frailejón, obtuvieron láminas con células superpuestas y donde la mayoría aún conservaba la lámina media.
En maíz, se reportó la obtención de buenos resultados utilizando hidrólisis ácida para la preparación de láminas citogenéticas (Chorzempa et al., 2017); sin embargo, esto no se observó en el presente estudio, lo cual podría deberse a varia-ciones en el material vegetal utilizado, el estado fisiológico del tejido o las condiciones experimen-tales particulares de cada procedimiento.
En cuanto a la hidrólisis enzimática, diversos estudios han reportado que el uso de mezclas enzimáticas genera preparaciones con células dispersas y en un solo plano (Lattier et al., 2017; Li et al., 2023). En maíz, McCaw et al. (2016) también obtuvieron buenos resultados empleando mezclas enzimáticas, además de señalar que el tiempo de digestión podía variar en función del tamaño del ápice radicular o de la edad de la enzima utilizada.
En nuestro estudio, la combinación de celulasa al 2% y pectinasa al 10% permitió obtener láminas con tinción adecuada, pero con abundante presencia de pequeñas vesículas tanto en el citoplasma como en el campo de observación. Estas estructuras podrían corresponder a restos de pared u orgánulos celulares producto de la degradación, o bien a oligosacáridos derivados de pectina generados durante la digestión enzimática (An et al., 2005; Pasarin et al., 2023), los cuales pueden agregarse y adoptar la apariencia de gránulos o vesículas.
Finalmente, respecto al tratamiento mediante hidrólisis combinada, se obtuvieron preparaciones limpias y con tinción uniforme. Estudios previos en diferentes especies vegetales también reportaron resultados satisfactorios con este tipo de hidrólisis (Delgado et al., 2010; Rodríguez & Bueno, 2006), señalando que los tratamientos ácidos o enzimáticos por separado resultan insuficientes (Rondón et al., 2013).
En este contexto, se resalta la importancia de aplicar el HCl tras el tratamiento enzimático en el proceso de preparación de láminas citogenéticas, dado su efecto en el aclaramiento del citoplasma y la remoción de vesículas y restos celulares que dificultan la visualización de las células al microscopio, así como en el ablandamiento del tejido para favorecer el posterior aplastamiento o squash. Una digestión adecuada de la pared celular, junto con una limpieza efectiva de restos celulares y maceración apropiada del tejido, mejora la penetración del colorante y explicaría la tinción nítida y uniforme resultante en las preparaciones.
Hora mitótica y ciclo celular
Los índices de fase parciales y el índice mitótico parcial se presentan en la Tabla 3. En Confite puntiagudo el mayor IM se registró a las 12:00 h (11,84 ± 2,18); sin embargo, no difirió significativamente de los valores obtenidos a las 11:00 h (8,15 ± 1,66) y 13:00 h (8,49 ± 1,41), aunque sí fue superior a los registrados a las 10:00, 14:00 y 15:00 h (Figura 3A). Por tanto, el intervalo de 11:00 a 13:00 h se considera el de mayor actividad mitótica para esta raza. En Blanco Urubamba, el máximo IM también se obtuvo a las 12:00 h (11,38 ± 1,03) y resultó significativamente mayor que en las demás horas evaluadas, por lo que la hora mitótica se ubica a las 12:00 h (Figura 3B). Por otro lado, Piricinco presentó el mayor IM a las 10:00 h (13,10 ± 4,45). No obstante, este valor no difirió significativamente de los registrados en las demás horas, lo que dificulta el establecimiento de una hora o rango definido de máxima actividad mitótica (Figura 3C).
En las tres razas, la fase del ciclo celular con mayor proporción de células correspondió a la interfase, mientras que, dentro de la mitosis, predominó la profase (Tabla 4), con una duración estimada de 0,27 h para Confite puntiagudo y Blanco Urubamba, y 0,28 h para Piricinco (Tabla 5). El porcentaje total de células en división (IMT) fue 8,51% en Confite puntiagudo, 8,39% en Blanco Urubamba y 9,79% en Piricinco (Tabla 4). En consecuencia, la duración del ciclo mitótico se estimó en aproximadamente 0,43 h para Confite puntiagudo, 0,42 h para Blanco Urubamba y 0,49 h para Piricinco (Tabla 5).
Tabla 3
Índices de fase parciales (IF) e índice mitótico parcial (IM) en ápices radiculares de tres razas peruanas de maíz evaluados entre las 10:00 y las 15:00 h
|
Raza |
Hora |
IFi (%) |
IFp (%) |
IFm (%) |
IFa (%) |
IFt (%) |
IM (%) |
|
CP |
10:00 |
93,59 ± 1,15 |
4,20 ± 0,90 |
1,32 ± 0,23 |
0,43 ± 0,12 |
0,46 ± 0,15 |
6,41 ± 1,15 |
|
11:00 |
91,85 ± 1,66 |
5,73 ± 2,06 |
1,53 ± 0,26 |
0,53 ± 0,30 |
0,37 ± 0,38 |
8,15 ± 1,66 |
|
|
12:00 |
88,16 ± 2,18 |
7,60 ± 2,17 |
2,28 ± 0,26 |
0,89 ± 0,55 |
1,06 ± 0,30 |
11,84 ± 2,18 |
|
|
13:00 |
91,51 ± 1,41 |
5,57 ± 1,58 |
1,56 ± 0,05 |
0,63 ± 0,06 |
0,73 ± 0,25 |
8,49 ± 1,41 |
|
|
14:00 |
92,22 ± 0,38 |
4,67 ± 0,57 |
1,46 ± 0,24 |
0,69 ± 0,17 |
0,96 ± 0,16 |
7,78 ± 0,38 |
|
|
15:00 |
92,23 ± 1,35 |
3,67 ± 1,88 |
1,52 ± 0,25 |
1,16 ± 0,29 |
1,42 ± 0,38 |
7,77 ± 1,35 |
|
|
BU |
10:00 |
92,00 ± 0,24 |
4,78 ± 0,81 |
1,26 ± 0,21 |
0,60 ± 0,10 |
1,36 ± 0,38 |
8,00 ± 0,24 |
|
11:00 |
91,66 ± 0,82 |
4,98 ± 0,57 |
1,83 ± 0,35 |
0,90 ± 0,36 |
0,63 ± 0,15 |
8,34 ± 0,82 |
|
|
12:00 |
88,62 ± 1,03 |
7,55 ± 1,11 |
2,05 ± 0,11 |
0,93 ± 0,30 |
0,86 ± 0,20 |
11,38 ± 1,03 |
|
|
13:00 |
91,81 ± 0,51 |
5,16 ± 0,70 |
1,60 ± 0,26 |
0,63 ± 0,25 |
0,80 ± 0,26 |
8,19 ± 0,51 |
|
|
14:00 |
93,97 ± 0,47 |
3,61 ± 0,25 |
1,09 ± 0,27 |
0,73 ± 0,16 |
0,60 ± 0,36 |
6,03 ± 0,47 |
|
|
15:00 |
91,58 ± 1,59 |
5,80 ± 0,81 |
1,49 ± 0,46 |
0,63 ± 0,57 |
0,50 ± 0,17 |
8,42 ± 1,59 |
|
|
PR |
10:00 |
86,90 ± 4,45 |
7,74 ± 2,99 |
2,45 ± 0,55 |
1,29 ± 0,65 |
1,62 ± 0,45 |
13,10 ± 4,45 |
|
11:00 |
88,81 ± 0,72 |
6,14 ± 0,38 |
2,49 ± 0,40 |
1,16 ± 0,46 |
1,40 ± 0,30 |
||
|
12:00 |
91,62 ± 2,34 |
4,44 ± 2,07 |
1,29 ± 0,11 |
0,73 ± 0,45 |
1,92 ± 0,40 |
8,38 ± 2,34 |
|
|
13:00 |
88,51 ± 2,30 |
7,38 ± 2,40 |
1,96 ± 0,21 |
1,26 ± 0,40 |
0,90 ± 0,43 |
11,49 ± 2,30 |
|
|
14:00 |
92,19 ± 1,46 |
4,51 ± 1,65 |
1,12 ± 0,15 |
0,47 ± 0,06 |
1,72 ± 0,49 |
7,81 ± 1,46 |
|
|
15:00 |
93,27 ± 1,42 |
3,33 ± 0,21 |
1,47 ± 0,55 |
0,67 ± 0,06 |
1,27 ± 0,84 |
6,73 ± 1,42 |
Los resultados se muestran en porcentajes (media ± desviación estándar). CP: Confite Puntiagudo, BU: Blanco Urubamba, PR: Piricinco, IF: índice de fase parcial, IM: índice mitótico parcial, i: interfase, p: profase, m: metafase, a: anafase y t: telófase.
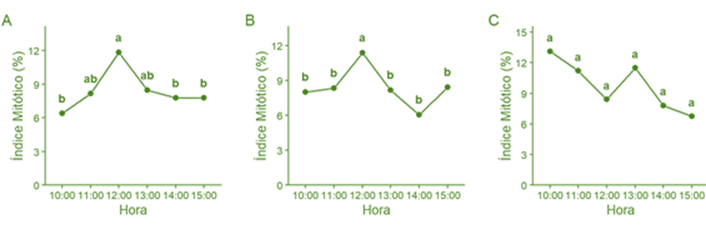
Figura 3. Variación del índice mitótico parcial en ápices radiculares de tres razas peruanas de maíz entre las 10:00 y 15:00 h. A. Confite puntiagudo B. Blanco Urubamba. C. Piricinco. Letras distintas indican diferencias significativas entre medias (p < 0,05).
Tabla 4
Índices de fase totales (IFT) e índice mitótico total (IMT) en ápices radiculares de tres razas peruanas de maíz
|
Raza |
IFTi (%) |
IFTp (%) |
IFTm (%) |
IFTa (%) |
IFTt (%) |
IMT (%) |
|
CP |
91,49 |
5,33 |
1,67 |
0,73 |
0,78 |
8,51 |
|
BU |
91,61 |
5,31 |
1,55 |
0,74 |
0,79 |
8,39 |
|
PR |
90,21 |
5,59 |
1,80 |
0,93 |
1,47 |
9,79 |
Los resultados se expresan en porcentajes. CP: Confite Puntiagudo, BU: Blanco Urubamba, PR: Piricinco, IFT: índice de fase total, IMT: índice mitótico total, i: interfase, p: profase, m: metafase, a: anafase y t: telofase.
Tabla 5
Duración estimada de las fases del ciclo celular (TF) y del ciclo mitótico (CM) en ápices radiculares de tres razas peruanas de maíz
|
Raza |
CC (h) |
TFi (h) |
TFp (h) |
TFm (h) |
TFa (h) |
TFt (h) |
CM (h) |
|
CP |
Tiempo de evaluación = 5h |
4,57 |
0,27 |
0,08 |
0,04 |
0,04 |
0,43 |
|
BU |
4,58 |
0,27 |
0,08 |
0,04 |
0,04 |
0,42 |
|
|
PR |
4,51 |
0,28 |
0,09 |
0,05 |
0,07 |
0,49 |
Los resultados se expresan en horas. CP: Confite Puntiagudo, BU: Blanco Urubamba, PR: Piricinco, CC: duración del ciclo celular, TF: duración de fase, i: interfase, p: profase, m: metafase, a: anafase, t: telofase y CM: duración del ciclo mitótico.
La determinación de la dinámica del ciclo celular y de la hora mitótica en especies vegetales constituye una herramienta ampliamente utilizada en una gran variedad de estudios citológicos (García-Godos Alcázar et al., 2020; Mata-Sucre et al., 2017; Quija-Lamina et al., 2010; Rodríguez & Bueno, 2006; Rondón et al., 2013). La hora mitótica se define como el momento del día en que el índice mitótico alcanza su valor máximo, indicando el periodo de mayor actividad de división celular. Su determinación permite reducir la influencia del azar en la elección del momento adecuado de muestreo (Talledo & Escobar, 1995).
En el presente estudio, la evaluación realizada entre las 10:00 a 15:00 h, permitió identificar un pico de actividad mitótica entre las 11:00 h a 13:00 h para Confite puntiagudo y alrededor del mediodía para Blanco Urubamba (Figura 3). De acuerdo con Alcorcés de Guerra (2002) la hora mitótica es un parámetro variable tanto entre especies como dentro de una misma especie, y suele ser característica de cada genotipo. Sin embargo, diferentes genotipos pueden compartir un mismo rango horario de máxima actividad mitótica. Deogade & Nasare (2016) reportaron que dos variedades de Allium sativum L. presentaban una hora mitótica similar entre las 9:00 y 10:00 h, lo que concuerda con los resultados obtenidos en este estudio, donde ambas razas de maíz mostraron un rango horario semejante de máxima actividad celular. Las horas mitóticas determinadas representarían los momentos más adecuados para la recolección de ápices radiculares en estudios citogenéticos, como la obtención de placas metafásicas para análisis cariotípico (Mata-Sucre et al., 2017; Rondón et al., 2013) o la determinación del número cromosómico (Quija-Lamina et al., 2010).
En contraste, en Piricinco no fue posible establecer una hora mitótica definida. Aunque a las 10:00 h se registró el valor máximo del índice mitótico parcial (IM), la elevada desviación estándar asociada—atribuida a la variabilidad entre réplicas—no permitió evidenciar diferencias significativas con respecto a las demás horas evaluadas (Figura 3).
Esta variabilidad podría atribuirse a múltiples factores. El índice mitótico es altamente sensible a factores ambientales extrínsecos como la intensidad lumínica (Miotto et al., 2021; Valencia Rincón & Rodríguez Castillo, 2012), la temperatura (Chorzempa et al., 2017; Neves et al., 2020), el estrés hídrico (Ribeiro et al., 2023; Rosely et al., 2024) o el estrés salino (Çavuşoğlu, 2023; Othman, 2020). Por ello, se ha empleado como indicador de citotoxicidad y tolerancia frente a diversos agentes físicos, químicos o ambientales (Bonea et al., 2018; El-Sagheer et al., 2024; Kassa, 2021; Murali et al., 2021).
Además de los factores ambientales, las características fenológicas propias de las semillas también influyen en el índice mitótico. Akhter et al. (1992) observaron que el envejecimiento y el almacenamiento prolongado de las semillas reducen el porcentaje de germinación y el índice mitótico en raíces de trigo y cebada. De forma similar, Dantas et al. (2019) documentaron una reducción del índice mitótico en raíces de semillas de arroz y frijol sometidas a envejecimiento artificial. En el caso del maíz, se ha señalado que la longevidad, las condiciones de almacenamiento y el momento de cosecha afectan el vigor de la semilla (Feng et al., 2024; Revilla et al., 2006).
En este sentido, considerando que la germinación, recolección de raíces, preparación de láminas y conteo celular se realizaron bajo las mismas condiciones experimentales, la elevada dispersión de los valores de IM en Piricinco podría explicarse por una posible heterogeneidad en la edad o vigor de las semillas utilizadas, aspecto que sería recomendable confirmar en evaluaciones posteriores aumentando el número de réplicas y verificando si la variabilidad se mantiene. Por otro lado, no debe descartarse la influencia de la información genética propia de cada raza, ya que, como señalan Anissa & Widodo (2021), la división celular varía entre organismos en función tanto de su composición genética como de factores ambientales (nutrición, temperatura, intensidad lumínica, etc.).
Respecto a los índices de fase totales (IFT), estos representan el porcentaje de células presentes en cada fase del ciclo celular y reflejan directamente la duración relativa de cada una de ellas (Rondón et al., 2013). Asimismo, el índice mitótico total (IMT) expresa la proporción de células en división con respecto al total de células observadas, reflejando la duración relativa de la mitosis o ciclo mitótico (CM).
Para estimar la duración de las fases y de la mitosis se consideró un valor promedio de duración del ciclo celular (CC) de 5 h—equivalente al intervalo de evaluación— el cual, de acuerdo con Talledo & Escobar (1995), corresponde a un tiempo suficiente en el que se observa claramente el inicio y el fin de la mitosis en un porcentaje significativo de células.
En las tres razas, la profase se identificó como la fase más prolongada y con mayor proporción de células en división dentro de la mitosis, mientras que la anafase presentó la menor frecuencia y duración (Tabla 5). Estos resultados concuerdan con lo reportado por Quispe et al. (2010), Rondón et al. (2013) y Wang et al. (1986), quienes también registraron a la profase como la etapa mitótica más extensa y a la anafase como la más breve. Sin embargo, algunos estudios no coinciden completamente con este patrón, evidenciando que la duración relativa de las fases mitóticas puede variar entre genotipos. Un ejemplo de esto fue descrito por Matos et al. (1995) y Valencia Rincón & Rodríguez Castillo (2012), quienes observaron en sus investigaciones que la telofase era la fase más corta del ciclo celular.
En el presente estudio, los valores de IMT obtenidos para Confite puntiagudo, Blanco Urubamba y Piricinco fueron de 8,51%, 8,39% y 9,79%, respectivamente. Como señala Dyer (1979, citado en Talledo & Escobar, 1995), el IMT no refleja la velocidad de división celular, sino la proporción de células en mitosis dentro del total en el ciclo celular, lo que permite estimar su duración para cada raza, que en nuestro caso se estimó en aproximadamente 0,43 h, para Confite puntiagudo, 0,42 h para Blanco Urubamba, y 0,49 para Piricinco (Tabla 5).
Una tendencia similar fue descrita por Verma & Lin (1979), quienes observaron en distintas líneas endogámicas de maíz una relativa constancia en la duración total del ciclo celular y la mitosis, atribuyendo esta estabilidad a la existencia de un control genético estricto y altamente específico que regula el ciclo celular. No obstante, la duración de la mitosis estimada por los autores fue mayor, entre 0,81 y 1,34 h, difiriendo de los valores obtenidos en el presente estudio. Esta diferencia se debería principalmente a que no establecieron una duración media del ciclo celular para estimar la duración de las fases, sino que calcularon directamente dichos valores mediante autorradiografía de células marcadas con timidina tritiada, estrategia que ha sido utilizada ampliamente en la comprensión de la cinética de la proliferación celular en meristemos radiculares (Desvoyes et al., 2021).
En contraste con la estabilidad antes descrita, diversos estudios han evidenciado que la dinámica del ciclo celular puede variar entre genotipos, asociándose a diferencias genéticas. Wang et al. (1986) reportaron que la duración del ciclo celular aumentaba en una variedad mexicana de maíz que presentaba cromosomas B frente a otra que no los presentaba, atribuyendo esta diferencia a que los cromosomas B prolongaban la fase S al darse su replicación de forma tardía durante dicho estadio. Asimismo, Jain & Gupta (1971) observaron diferencias en la duración del ciclo celular—particularmente en la fase S—entre variedades primitivas y mejoradas de maíz, sugiriendo que las variedades primitivas presentaban un mayor contenido de ADN en sus células en comparación con las variedades más evolucionadas.
Por otra parte, se ha demostrado que estas variaciones pueden originarse a partir de diferencias en los mecanismos de regulación génica. Huang et al. (2019) identificaron en el maíz el gen Vks1 como un regulador de la mitosis y citocinesis durante el desarrollo temprano del endospermo, y propusieron que las alteraciones en su regulación podrían provocar una represión de la división celular. Por su parte, Zhao et al. (2022) demostraron que los genes de las ciclinas no solo regulan la progresión del ciclo celular, sino como es el caso del ZmCYCB1-1, también regulan el crecimiento del embrión y tamaño de la semilla.
Finalmente, diversos factores ambientales, especialmente los de tipo abiótico, pueden influir en la duración del ciclo celular. Zhao et al. (2014) reportaron que distintas condiciones de estrés abiótico, como la salinidad y las variaciones extremas de temperatura, incrementan los niveles de acetilación de genes específicos del ciclo celular, prolongando su duración y afectando el crecimiento y desarrollo de las plántulas de maíz.
De igual manera, Qi & Zhang (2020) señalaron que factores abióticos como la salinidad, la temperatura, la humedad y la luz influyen significativamente en el crecimiento de las plantas y el rendimiento de los cultivos, efectos que se atribuyen a alteraciones en los procesos de proliferación y expansión celular. En este mismo contexto, Hodgkinson et al. (2023) refieren que los procesos de estrés por factores físicos, metabólicos o químicos pueden producir daños en el ADN y comprometer el ciclo celular, y por ello es fundamental investigar los parámetros de la dinámica celular.
En conjunto, los resultados sugieren que, si bien las diferencias en la duración del ciclo mitótico y de sus fases fueron mínimas entre las tres razas de maíz estudiadas, factores genéticos y ambientales, así como variaciones en los mecanismos de regulación génica, pueden influir en la dinámica del ciclo celular. Chu & Wang (2023) destacan que uno de los principales desafíos actuales es comprender el mecanismo del ciclo celular desde la perspectiva estructural cromosómica dado que los cromosomas presentan una dinámica propia a lo largo de las distintas fases del ciclo celular.
Si bien el análisis de la dinámica estructural cromosómica no fue abordado directamente, los resultados aquí presentados contribuyen a cubrir un primer eslabón en el conocimiento de la dinámica del ciclo celular y determinación de la hora mitótica en maíz peruano, lo que permite proyectar investigaciones posteriores orientadas al análisis integrado de la dinámica celular y cromosómica. En este sentido, el presente estudio se sustenta en análisis citológicos que permiten evaluar de manera práctica y eficiente la dinámica del ciclo celular en maíz y establecer su hora mitótica; información que resulta fundamental por su aplicabilidad en estudios futuros de mayor alcance y complejidad en el campo de la citogenética vegetal.
CONCLUSIONES
Para el estudio de la hora mitótica y del ciclo celular en tres razas peruanas de maíz se realizó la estandarización del protocolo de preparación de láminas citogenéticas a partir de meristemos apicales radiculares. En el paso crítico de hidrólisis, el mejor tratamiento correspondió a la hidrólisis enzimática seguida de hidrólisis ácida, obtenién-dose láminas limpias y de tinción uniforme. La hora mitótica se estimó alrededor del mediodía para Confite puntiagudo y Cuzco gigante var. Blanco Urubamba, mientras que en Piricinco no se definió un rango horario específico de máxima actividad mitótica, posiblemente debido a características fisiológicas propias de las semillas, aspecto que sería importante evaluar en futuras investi-gaciones. La duración del ciclo mitótico fue similar entre razas, oscilando entre 0,42 h y 0,49 h, lo que sugiere una posible conservación en la dinámica del ciclo celular. En perspectiva, la información obtenida sobre la dinámica del ciclo celular y la determinación de la hora mitótica en razas peruanas de maíz constituye una base método-lógica para el desarrollo de estudios citogenéticos de mayor alcance, que incluyan un mayor número de razas y la evaluación bajo distintas condiciones ambientales. Los datos obtenidos posibilitarán optimizar estudios citogenéticos de mayor complejidad que involucren mapeo físico de cromosomas y análisis cariotípico detallado, permitiendo realizar investigaciones orientadas al análisis de la estructura y dinámica cromosómica, identificación de haploidías, relaciones evolutivas y/o estudios comparativos de genomas. Asimismo, estos aportes pueden fortalecer su aplicación en programas de mejoramiento genético y conservación del germoplasma nativo.
AGRADECIMIENTOS
Agradecemos al Vicerrectorado de Investigación y Posgrado de la UNMSM por el financiamiento del proyecto de investigación B22100631 (RR 005557-2022-R/UNMSM), producto del cual deriva la presente publicación. Asimismo, agradecemos al Banco de Germoplasma del Programa de Investigación y Proyección Social en Maíz de la UNALM, en especial al Ingeniero Gean Carlo Ciprian Salcedo, por la donación del material biológico utilizado.
REFERENCIAS BIBLIOGRÁFICAS
Aguilera, P. M., Debat, H. J., Scaldaferro, M. A., Martí, D. A., & Grabiele, M. (2016). FISH-mapping of the 5S rDNA locus in chili peppers (Capsicum-Solanaceae). Anais da Academia Brasileira de Ciências, 88(1), 117–125. https://doi.org/10.1590/0001-37652301620140616
Akhter, F. N., Kabir, G., Mannan, M. A., & Shaheen, N. (1992). Aging effect of wheat and barley seeds upon germination, mitotic index and chromosomal damage. Medical Journal of Islamic World Academy of Sciences, 5(1), 44–48.
Alcorcés de Guerra, N. (2002). Cariología de dos especies del género Tabebuia Gomes (Bignoniaceae). Revista Científica UDO Agrícola, 2(1), 14–21.
Aliyeva-Schnorr, L., Ma, L., & Houben, A. (2015). A fast air-dry dropping chromosome preparation method suitable for FISH in plants. Journal of Visualized Experiments, 106, 53470. https://doi.org/10.3791/53470
An, H. J., Lurie, S., Greve, L. C., Rosenquist, D., Kirmiz, C., Labavitch, J. M., & Lebrilla, C. B. (2005). Determination of pathogen-related enzyme action by mass spectrometry analysis of pectin breakdown products of plant cell walls. Analytical Biochemistry, 338(1), 71–82. https://doi.org/10.1016/j.ab.2004.11.004
Annisa, S., & Widodo. (2021). Studies on mitotic division of Allium ascalonicum L. based on observation time. Proceeding of the International Conference on Science and Engineering, 4, 53–55.
Arias Salcedo, R. V. F., Chávez Santa Cruz, G., & Medina Hinostroza, T. (2025). Bioseguridad y conservación de la biodiversidad del maíz (Zea mays L.) en Perú. Manglar, 22(1), 123-134. http://doi.org/10.57188/manglar.2025.014
Barrera-Guzmán, L. A., Legaria-Solano, J. P., & Ortega-Paczka, R. (2020). Diversidad genética en poblaciones de razas mexicanas de maíz. Revista Fitotecnia Mexicana, 43(1), 121–125. https://doi.org/10.35196/rfm.2020.1.121
Bian, J., Zhuang, Z., Tang, R., Ta, W., Ren, Z., & Peng, Y. (2025). Single-cell RNA sequencing reveals cellular diversity and gene expression dynamics in maize root development. Frontiers in Plant Science, 16, 1666531. https://doi.org/10.3389/fpls.2025.1666531
Bonea, D., Bonciu, E., Niculescu, M., & Olaru, A. L. (2018). The allelopathic, cytotoxic and genotoxic effect of Ambrosia artemisiifolia on the germination and root meristems of Zea mays. Caryologia, 71(1), 24–28. https://doi.org/10.1080/00087114.2017.1400263
Burbano, L., Fernández, E., & Cortés, E. (2011). Determinación de la hora mitótica de Guadua angustifolia var. grandicaula (Familia: Gramineae) a partir de meristemos radiculares. Revista de Investigaciones Universidad del Quindío, 22(1), 129–135. https://doi.org/10.33975/riuq.vol22n1.667
Çavuşoğlu, D. (2023). Modulation of NaCl-induced osmotic, cytogenetic, oxidative and anatomic damages by coronatine treatment in onion (Allium cepa L.). Scientific Reports, 13(1), 1580. https://doi.org/10.1038/s41598-023-28849-w
Cheng, J., & Wang, W. (2025). Meta-analysis of genetic diversity in global fresh-eating maize germplasm resources. Maize Genomics and Genetics, 16(2). https://doi.org/10.5376/mgg.2025.16.0007
Chorzempa, S. E., Perniola, O. S., Moscheñski, S. M., López, C., Pantuso, F., & Molina, M. del C. (2017). El índice mitótico como parámetro estimador de la tolerancia a bajas temperaturas durante la germinación de Zea mays. Revista de la Facultad de Agronomía, 116(2), 259–266.
Chu, X., & Wang, J. (2023). Quantifying the large-scale chromosome structural dynamics during the mitosis-to-G1 phase transition of the cell cycle. Open Biology, 13(11), 230175. https://doi.org/10.1098/rsob.230175
Dantas, A. F., Fascineli, M. L., José, S. C. B. R., Pádua, J. G., Gimenes, M. A., & Grisolia, C. K. (2019). Loss of genetic integrity in artificially aged seed lots of rice (Oryza sativa L.) and common bean (Phaseolus vulgaris L.). Mutation Research: Genetic Toxicology and Environmental Mutagenesis, 846, 503–509. https://doi.org/10.1016/j.mrgentox.2019.07.008
Delgado, L. M., Uribe Lastra, M., & Marulanda Ángel, M. L. (2010). Estandarización de la técnica citogenética “squash” para conteo de cromosomas mitóticos en Rubus glaucus Benth. Scientia et Technica, 17(46), 74–79.
Deogade, S., & Nasare, P. N. (2016). Mitotic index (MI) and phase index (PI) in Allium sativum Linn. (varieties Agrifound White and Yamuna Safed). International Journal of Recent Biotechnology and Applied Technology, 4(1), 188–192.
Desvoyes, B., Echevarría, C., & Gutierrez, C. (2021). A perspective on cell proliferation kinetics in the root apical meristem. Journal of Experimental Botany, 72(19), 6708–6721. https://doi.org/10.1093/jxb/erab303
Domínguez, P. G., Freilij, D., & Lia, V. V. (2024). A genomic journey across the past, present, and future of South American maize. Journal of Experimental Botany, 75(22), 7006–7018. https://doi.org/10.1093/jxb/erae383
Echevarría, C., Gutiérrez, C., & Desvoyes, B. (2021). Tools for assessing cell-cycle progression in plants. Plant and Cell Physiology, 62(8), 1231–1238. https://doi.org/10.1093/pcp/pcab066
El-Sagheer, S. M., Ahmed, S. A., Abdu-Allah, G. A. M., Ali, A. H., & Ghanem, A.-M. F. M. (2024). Cytogenetical impact of some herbicides and their combinations on maize (Zea mays L.) seedling root tip cells. Journal of Agricultural Sciences and Engineering, 6(1), 32–37. https://doi.org/10.48309/jase.2024.187128
Feng, W., Wang, H., Bai, Y., Wang, W., Xue, W., Shi, Z., Qiu, P., Gao, Y., Xue, J., Sun, M., & Ma, H. (2024). Phenotypic, physiological and hormonal analysis reveals the mechanisms of timely harvesting for ensuring the seed vigor of maize (Zea mays L.) inbred lines. Agronomy, 14(8), 1770. https://doi.org/10.3390/agronomy14081770
Ferrer Pereira, H. E., Alcorcés de Guerra, N., & Méndez-Natera, J. (2007). Determinación del ciclo mitótico de dos cultivares de Gossypium hirsutum L. y dos ecotipos de Gossypium barbadense L. Acta Biológica Paranaense, 36(3–4), 121–149.
Fuentes-Cardenas, I. S., Cuba-Puma, R., Marcilla-Truyenque, S., Begazo-Gutiérrez, H., Zolla, G., Fuentealba, C., Shetty, K., & Ranilla, L. G. (2022). Diversity of the Peruvian Andean maize (Zea mays L.) race Cabanita: Polyphenols, carotenoids, in vitro antioxidant capacity, and physical characteristics. Frontiers in Nutrition, 9, 983208. https://doi.org/10.3389/fnut.2022.983208
García Mendoza, P. J. (2017). El cultivo del maíz en el mundo y en Perú. Revista de Investigaciones de la Universidad Le Cordon Bleu, 4(2), 73–79. https://doi.org/10.36955/riulcb.2017v4n2.005
García-Godos Alcázar, P., Palomino Felices, S., & Martínez Gómez, K. (2020). Diversidad citogenética de Physalis peruviana L. “aguaymanto” de los ecotipos del Perú. Investigación, 28(1), 157–165. https://doi.org/10.51440/unsch.revistainvestigacion.28.1.2020.368
Hake, S., & Ross-Ibarra, J. (2015). Genetic, evolutionary and plant breeding insights from the domestication of maize. eLife, 4, e05861. https://doi.org/10.7554/eLife.05861
Harun, A., Liu, H., Song, S., Asghar, S., Wen, X., Fang, Z., & Chen, C. (2023). Oligonucleotide fluorescence in situ hybridization: An efficient chromosome painting method in plants. Plants, 12(15), 2816. https://doi.org/10.3390/plants12152816
Hodgkinson, A., Tursynkozha, A., & Trucu, D. (2023). Structured dynamics of the cell-cycle at multiple scales. Frontiers in Applied Mathematics and Statistics, 9, Article 1090753. https://doi.org/10.3389/fams.2023.1090753
Huang, Y., Wang, H., Huang, X., Wang, Q., Wang, J., An, D., Li, J., Wang, W., & Wu, Y. (2019). Maize VKS1 regulates mitosis and cytokinesis during early endosperm development. The Plant Cell, 31(6), 1238–1255. https://doi.org/10.1105/tpc.18.00966
Jaimes Méndez, N., Gil Durán, M. A., & Jaimes Méndez, D. A. (2022). Estudio comparativo del índice mitótico en el meristemo de Allium cepa L. Bistua. Revista de la Facultad de Ciencias Básicas, 20(1), 30–33. https://doi.org/10.24054/bistua.v20i1.1158
Jain, K., & Gupta, P. K. (1971). Duration of mitotic cycle in evolved and primitive varieties of maize. Maize Genetics Cooperation Newsletter, 45, 53–57.
Kaduchová, K., Marchetti, C., Ovečka, M., Galuszka, P., Bergougnoux, V., Šamaj, J., & Pecinka, A. (2023). Spatial organization and dynamics of chromosomes and microtubules during barley mitosis. The Plant Journal, 115(3), 602–613. https://doi.org/10.1111/tpj.16355
Kassa, B. A. (2021). Cytotoxicity and genotoxicity evaluation of municipal wastewater discharged into the head of Blue Nile River using the Allium cepa test. Scientific African, 13, e00911. https://doi.org/10.1016/j.sciaf.2021.e00911
Kawano, S. (1965). Application of pectinase and cellulase in an orcein squash method. Shokubutsugaku Zasshi, 78(919), 36–42. https://doi.org/10.15281/jplantres1887.78.36
Khalaf, S. D., Alhamadany, A. Y. M., & Kader, M. A. (2023). The fluorescence in situ hybridization (FISH): Types and applications—A review. NTU Journal of Pure Sciences, 2(2), 38–48. https://doi.org/10.56286/ntujps.v2i2.416
Kim, K.-H., Park, D., & Lee, B.-M. (2026). Biotechnological strategies to enhance maize resilience under climate change. Biology, 15(2), 161. https://doi.org/10.3390/biology15020161
Lattier, J. D., Chen, H., & Contreras, R. N. (2017). Improved method of enzyme digestion for root tip cytology. HortScience, 52(7), 1029–1032. https://doi.org/10.21273/hortsci12024-17
Li, L., Schwarzacher, T., Tomaszewska, P., Liu, Q., Li, X. Z., Yi, K., Wu, W., & Heslop-Harrison, J. S. (P.) (2023). Protocols for chromosome preparations: Molecular cytogenetics and studying genome organization in coffee. In I. L. W. Ingelbrecht, F. A. da Silva, & N. Jankowicz-Cieslak (Eds.), Mutation breeding in coffee with special reference to leaf rust: Protocols (pp. 291–314). Springer. https://doi.org/10.1007/978-3-662-67273-0_21
Liu, J., Fernie, A. R., & Yan, J. (2020). The past, present, and future of maize improvement: Domestication, genomics, and functional genomic routes toward crop enhancement. Plant Communications, 1(1), 100010. https://doi.org/10.1016/j.xplc.2019.100010
Mata-Sucre, J., Imery-Buzia, J., & Cequea-Ruíz, H. (2017). Estudio cariotípico en plantas de Agave cocui Trelease (Asparagaceae) nativas del estado Sucre, Venezuela. Agrobiología, 29(0), 174–182.
Matos, Á., Molina, J., Acosta, D., & Abreu, M. (1995). Determinación del proceso mitótico en células radicales de Prosopis juliflora DC. Ciencia, 3(2), 83–90.
McCaw, M., Swyers, N., Graham, N., Cody, J., Zhao, C., & Birchler, J. (2016). Preparation of chromosomes from Zea mays. Current Protocols in Plant Biology, 1(3), 501–509. https://doi.org/10.1002/cppb.20032
Ministerio del Ambiente (MINAM). (2018). Línea de base de la diversidad genética del maíz peruano con fines de bioseguridad (T. M. Hinostroza, D. Cañedo Torres, C. Aguirre Asturrizaga, & H. Tello Fernández, Eds., 1.ª ed.) Grupo Raso.
Miotto, Y. E., da Costa, C. T., Offringa, R., Kleine-Vehn, J., & Maraschin, F. dos S. (2021). Effects of light intensity on root development in a D-Root growth system. Frontiers in Plant Science, 12, 778382. https://doi.org/10.3389/fpls.2021.778382
Mondin, M., Santos-Serejo, J. A., Bertão, M. R., Laborda, P., Pizzaia, D., & Aguiar-Perecin, M. L. R. (2014). Karyotype variability in tropical maize sister inbred lines and hybrids compared with KYS standard line. Frontiers in Plant Science, 5, 544. https://doi.org/10.3389/fpls.2014.00544
Murali, M., Anandan, S., Ansari, M. A., Alzohairy, M. A., Alomary, M. N., Asiri, S. M. M., Almatroudi, A., Thriveni, M. C., Singh, S. B., Gowtham, H. G., Aiyaz, M., Srinivasa, C., Urooj, A., & Amruthesh, K. N. (2021). Genotoxic and cytotoxic properties of zinc oxide nanoparticles phyto-fabricated from the obscure morning glory plant Ipomoea obscura (L.) Ker Gawl. Molecules, 26(4), 891. https://doi.org/10.3390/molecules26040891
Neves, V., Viegas, W., & Caperta, A. D. (2020). Effects of high temperature on mitotic index, microtubule and chromatin organization in rye (Secale cereale L.) root-tip cells. Caryologia, 73(4), 55–63. https://doi.org/10.13128/caryologia-788
Othman, H. (2020). Impact of different concentrations of sodium chloride on root growth, cell division and chromosomal abnormalities in the root tips of Allium cepa. Journal of Advanced Laboratory Research in Biology, 11(1), 1–6.
Palacios-Rojas, N., McCulley, L., Kaeppler, M., Titcomb, T. J., Gunaratna, N. S., Lopez-Ridaura, S., & Tanumihardjo, S. A. (2020). Mining maize diversity and improving its nutritional aspects within agro-food systems. Comprehensive Reviews in Food Science and Food Safety, 19(4), 1809–1834. https://doi.org/10.1111/1541-4337.12552
Panda, S., Borkataki, S., Reddy, M. D., & Nanda, S. P. (2020). Domestication and evolution of maize. International Journal of Chemical Studies, 8(1), 1834–1838. https://doi.org/10.22271/chemi.2020.v8.i1aa.8530
Pasarin, D., Ghizdareanu, A. I., Teodorescu, F., Rovinaru, C., & Banu, A. (2023). Characterization of pectin oligosaccharides obtained from citrus peel pectin. Fermentation, 9(3), 312. https://doi.org/10.3390/fermentation9030312
Qi, F., & Zhang, F. (2020). Cell cycle regulation in the plant response to stress. Frontiers in Plant Science, 10, 1765. https://doi.org/10.3389/fpls.2019.01765
Quija-Lamina, P., Segovia-Salcedo, C., Jadán, M., & Proaño, K. (2010). Estandarización de la metodología para el conteo cromosómico en especies del género Polylepis en el Ecuador. Revista Ecuatoriana de Medicina y Ciencias Biológicas, 31(1–2). https://doi.org/10.26807/remcb.v31i1-2.41
Quispe, J., Saldaña, J., Verde, T., & Valderrama, S. (2010). Efectos del sorbato de potasio a diferentes concentraciones y tiempo de exposición sobre el ciclo celular y el material genético en meristemos radiculares de Allium cepa L. Revista del Encuentro Científico Internacional, 7(1). https://doi.org/10.33017/RevECIPeru2010.0011/
Revilla, P., Velasco, P., Malvar, R. A., Cartea, M. E., & Ordás, A. (2006). Variability among maize (Zea mays L.) inbred lines for seed longevity. Genetic Resources and Crop Evolution, 53(4), 771–777. https://doi.org/10.1007/s10722-004-5542-1
Ribeiro, S., Gaspar, M. J., Lima-Brito, J., Fonseca, T., Soares, P., Cerveira, A., Fernandes, P. M., Louzada, J., & Carvalho, A. (2023). Impact of fire recurrence and induced water stress on seed germination and root mitotic cell cycle of Pinus pinaster Aiton. Forests, 14(1), 78. https://doi.org/10.3390/f14010078
Rodríguez, N. C., & Bueno, M. L. (2006). Estudio de la diversidad citogenética de Physalis peruviana L. (Solanaceae). Acta Biológica Colombiana, 11(2), 75–85.
Rondón, H. B., Rache, L. C., & Pacheco, J. M. (2013). Cariotipo de Espeletiopsis muiska. Revista MVZ Córdoba, 18(3), 3868–3876.
Rosely, N. F. S., Saimi, N. N., Midin, M. R., & Karim, M. F. (2024). Acclimation to drought stress improves root physiology and cell mitotic index, leaf pigments and water status in Oryza sativa L. International Journal of Biology and Chemistry, 17(2), 14–24. https://doi.org/10.26577/ijbch2024v17.i2.2
Salhuana, W. S. (2003). Diversidad y descripción de las razas de maíz en el Perú (pp. 204–251). En W. Salhuana, A. Valdez, F. Scheuch, & J. Davelouis (Eds.), Programa Cooperativo de Investigaciones en Maíz (PCIM): Logros y perspectivas, 50° aniversario. Universidad Nacional Agraria La Molina. Lima, Perú.
Salvador-Reyes, R., & Clerici, M. T. P. S. (2020). Peruvian Andean maize: General characteristics, nutritional properties, bioactive compounds, and culinary uses. Food Research International, 130, 108934. https://doi.org/10.1016/j.foodres.2019.108934
Sanodiya, P., Tiwari, P., & Gupta, C. (2023). Maize (Zea mays L.): The future potential cereal crop of indispensable significance in agroforestry systems. Maize Journal, 12(2), 61–71.
Sauer, D. B., & Burroughs, R. (1986). Disinfection of seed surfaces with sodium hypochlorite. Phytopathology, 76(7), 745. https://doi.org/10.1094/phyto-76-745
Song, Y., Wang, J., Ma, Y., Wang, J., Bao, L., Sun, D., Lin, H., Fan, J., Zhou, Y., Zeng, X., Wang, Z., Zhang, L., Li, C., & Di, H. (2025). Genetic Diversity and Construction of Salt-Tolerant Core Germplasm in Maize (Zea mays L.) Based on Phenotypic Traits and SNP Markers. Plants, 14 (14), 2182. https://doi.org/10.3390/plants14142182
Talledo, D., & Escobar, C. (1995). El ciclo celular en vegetales: Su estudio, importancia y aplicaciones. Biotiempo, 2, 13–31. https://doi.org/10.31381/biotiempo.v2i0.1531
Torres-García, S. R., Rojas-Vásquez, Z., Baselly-Villanueva, J. R., Salazar-Ramos, I., Terán-Piña, J. C., Gutiérrez-Arce, F. B., & Roque-Alcarraz, R. E. (2025). Caracterización de la calidad nutricional de 10 variedades de maíz con destino forrajero en Tarapoto. Revista de Veterinaria y Zootecnia Amazónica, 5(2), e1150. https://doi.org/10.51252/revza.v5i2.1150
Valencia-Rincón, J. C., & Rodríguez-Castillo, N. (2012). Estimación de la duración del ciclo celular y estandarización del protocolo citogenético en Guadua angustifolia Kunth var. angustifolia (Bambusoideae, Poaceae). Revista de Investigaciones Universidad del Quindío, 23(2), 81–91. https://doi.org/10.33975/riuqu.vol23n2.407
Valladolid, A., Blas, R., & Gonzáles, R. (2004). Introducción al recuento de cromosomas somáticos en raíces andinas. En J. Seminario (Ed.), Raíces andinas: Contribuciones al conocimiento y la capacitación (Serie Conservación y uso de la biodiversidad de raíces y tubérculos andinos: Una década de investigaciones para el desarrollo [1993–2003], 6, pp. 96–99). Universidad Nacional de Cajamarca; Centro Internacional de la Papa; Agencia Suiza para el Desarrollo y la Cooperación. Lima, Perú.
Verma, R. S., & Lin, M. S. (1979). The duration of DNA synthetic (S) period in Zea mays: A genetic control. Theoretical and Applied Genetics, 54(6), 277–282. https://doi.org/10.1007/bf00281210
Wang, A. S., Phillips, R. L., & Mi, C. C. (1986). Cell cycle parameters and accumulation of metaphase cells in maize suspension cultures. Plant Science, 46(1), 53–61. https://doi.org/10.1016/0168-9452(86)90131-7
Yan, J., & Tan, B. C. (2019). Maize biology: From functional genomics to breeding application. Journal of Integrative Plant Biology, 61(6), 654–657. https://doi.org/10.1111/jipb.12819
Yao, Q., Gao, J., Chen, F., & Li, W. (2020). Development and application of an optimized drop-slide technique for metaphase chromosome spreads in maize. Biotechnic & Histochemistry, 95(4), 276–284. https://doi.org/10.1080/10520295.2019.1686167
Zambrano Mendoza, J. L., Albán Guijarro, M. G., & Caviedes Cepeda, G. M. (2025). Resultados e impactos de la gestión del conocimiento en el desarrollo de tecnologías para el cultivo de maíz en las Américas. Bitácora Académica, 16. https://doi.org/10.18272/5wtr7w70
Zhao, L., Wang, P., Hou, H., Zhang, H., Wang, Y., Yan, S., Huang, Y., Li, H., Tan, J., Hu, A., Gao, F., Zhang, Q., Li, Y., Zhou, H., Zhang, W., & Li, L. (2014). Transcriptional regulation of cell cycle genes in response to abiotic stresses correlates with dynamic changes in histone modifications in maize. PLoS ONE, 9(8), e106070. https://doi.org/10.1371/journal.pone.0106070
Zhao, B., Zhou, M., Ren, W., Li, H., Zhang, Q., He, G., Liu, Y., & He, H. (2022). The B-type cyclin CYCB1-1 regulates embryonic development and seed size in maize. International Journal of Molecular Sciences, 23(11), 5907. https://doi.org/10.3390/ijms23115907