 |
INTRODUCCIÓN
Detección de OVM en maíz nativo: Discrepancias entre métodos inmunocromatográficos y moleculares en mercados de Lima, Perú
Detection of LMOs in native maize: Discrepancies between immunochromatographic and molecular methods in markets in Lima, Peru
Tiffany Aylyn Tinoco Calizaya+1, 2, 3; Ruby Danixa Gomez Espinoza+1, 2, 3; David Eduardo Castro Garro3, 4; Jorge Augusto Sanchez Ayte5; Ruben Armando Daga Lopez1, 2, 3; Zanhy Leonor Valencia Reyes2, 3;
Jacinto Joaquín Vertiz Osores1, 2, 3; Obert Marín-Sánchez1, 2, 3*
1 Programa de Maestría en Biotecnología y Gestión Ambiental. Universidad Nacional Tecnológica de Lima Sur. Ciudad Universitaria, Av. Bolivar S/N, Sector 3 Grupo 1, Mz. A, Sublote 3, Villa El Salvador, Lima, Perú.
2 Laboratorio de Investigación “Evaluación Ambiental y Diversidad Biológica” (EVADIB). Universidad Nacional Tecnológica de Lima Sur. Ciudad Universitaria, Av. Bolivar S/N, Sector 3 Grupo 1, Mz. A, Sublote 3, Villa El Salvador, Lima, Perú.
3 Grupo de Investigación Bioprospección en Salud y Metagenómica Ambiental – BIOPROSGEN. Universidad Nacional Tecnológica de Lima Sur. Ciudad Universitaria, Av. Bolivar S/N, Sector 3 Grupo 1, Mz. A, Sublote 3, Villa El Salvador, Lima, Perú.
4 Dirección de Recursos Genéticos y Bioseguridad. Ministerio del Ambiente, Av. Antonio Miró Quesada 425, Magdalena del Mar, Lima, Perú.
5 Grupo de Investigación Innovación Tecnológica Multidisciplinaria en Ciencias e Ingeniería – MITSE. Universidad Nacional Tecnológica de Lima Sur. Ciudad Universitaria, Av. Bolivar S/N, Sector 3 Grupo 1, Mz. A, Sublote 3, Villa El Salvador, Lima, Perú.
+ Igual contribución en la primera autoría.
* Autor corresponsal: omarins@untels.edu.pe (O. Marín-Sánchez).
ORCID de los autores:
T. A. Tinoco Calizaya: https://orcid.org/0009-0001-9218-5211 R. D. Gomez Espinoza: https://orcid.org/0009-0000-3530-8273
D. E. Castro Garro: https://orcid.org/0000-0003-3182-1744 J. A. Sánchez Ayte: https://orcid.org/0000-0001-9734-3381
R. A. Daga Lopez: https://orcid.org/0000-0002-3105-1594 Z. L. Valencia Reyes: https://orcid.org/0000-0003-0271-7104
J. J. Vértiz Osores: https://orcid.org/0000-0003-2774-1207 O. Marín-Sánchez: https://orcid.org/0000-0003-2912-1191
RESUMEN
El objetivo fue detectar la presencia de organismos vivos modificados (OVM) en maíz (Zea mays), de las variedades nativas “chullpi” y “morado”, vendidos a granel en los mercados de Lima Sur. Se realizó un muestreo en 30 mercados (60 muestras). Se emplearon tiras inmunocromatográficas (QuickComb®) para un tamizaje de proteínas transgénicas y se confirmaron los resultados con PCR en tiempo real (rt-PCR) para detectar el Promotor-35S y Terminador-NOS. Como resultado, las pruebas QuickComb® detectaron nueve (9) proteínas transgénicas en ambos tipos de maíz. Sin embargo, todos los resultados de rt-PCR fueron negativos para las secuencias P-35S y T-NOS. Se concluye que los positivos en las tiras reactivas pueden ser falsos positivos, probablemente causados por contaminación cruzada superficial en el maíz “chullpi” y por interferencia no específica (posiblemente por antocianinas) en el maíz “morado”. El estudio subraya la necesidad de confirmación molecular para la vigilancia de OVM en granos de maíces nativos.
Palabras clave: OVM; maíz nativo; falsos positivos; PCR en tiempo real; bioseguridad.
ABSTRACT
The objective was to detect the presence of living modified organisms (LMOs) in “chullpi” and “morado” native landraces of maize (Zea mays) sold in bulk in markets in Southern Lima. Sampling was conducted in 30 markets (60 samples). Immunochromatographic strips (QuickComb®) were used for screening transgenic proteins, and the results were confirmed with real-time PCR (rt-PCR) to detect the 35S Promoter and NOS Terminator. As a result, the QuickComb® tests detected nine (9) transgenic proteins in both types of maize. However, all RT-PCR results were negative for the P-35S and T-NOS sequences. It is concluded that the positive results on the reagent strips may be false positives, probably caused by surface cross-contamination in the “chullpi” maize and by non-specific interference (possibly from anthocyanins) in the “morado” maize. The study underscores the need for molecular confirmation for LMO surveillance in grain native maize landraces.
Keywords: LMO; native corn; false positives; real-time PCR; biosecurity.
Recibido: 28-01-2026.
Aceptado: 14-03-2026.
|
El Perú es un centro de diversificación primaria y de domesticación secundaria del maíz (Zea mays L.), albergando una vasta agrobiodiversidad que es pilar de la seguridad alimentaria y de la cultura andina (Salvador-Reyes & Clerici, 2020). La preservación de esta riqueza genética frente a la potencial introgresión de Organismos Vivos Modificados (OVM) es una prioridad nacional (MINAM, 2018) no solo por el riesgo de la pérdida del recurso biológico per se, sino por la potencial vulnerabilidad de la especie frente al cambio climático (Zheng et al., 2024). Variedades nativas como el maíz “chullpi” y el maíz “morado”, comercializadas ampliamente en mercados locales, son vulnerables a la coexistencia y eventual flujo genético con maíces transgénicos, lo que representa riesgos ecológicos, económicos y sociales que deben ser evaluados, tal como lo establece el Protocolo de Cartagena sobre Seguridad de la Biotecnología del Convenio sobre la Diversidad Biológica (Cartagena, 2020).
En respuesta a esta amenaza, el marco legal peruano ha establecido mecanismos de bioseguridad. El más significativo es la Ley N.º 29811, extendida por la Ley N.º 31111 (Congreso de la República del Perú, 2021), que establece una moratoria al ingreso y producción de OVM con fines de cultivo en el territorio nacional hasta el año 2035. Esta medida de precaución busca fortalecer las capacidades nacionales de investigación y bioseguridad antes de tomar decisiones sobre la liberación de estos organismos. Esta norma establece un sistema de control de las importaciones y monitoreos en campo para detectar el ingreso o liberación ilegal de OVM en el ambiente. Pero, a pesar de esta restricción, se tienen reportes formales de ingresos clandestinos de semillas modificadas a los campos de cultivo del territorio nacional, que podría transformarse en un problema invisibilizado requiriéndose de la atención gubernamental, a pesar de los beneficios del cultivo indicados en la literatura (Naqvi et al., 2011).
Paralelamente, el Código de Protección y Defensa del Consumidor peruano (Ley N.º 29571) (Congreso de la República del Perú, 2010), estipula el derecho de los consumidores a ser informados sobre el contenido de los alimentos, que se encuentra a disposición de compra, incluyendo la presencia de componentes OVM. No obstante, la efectividad de estas regulaciones depende de un monitoreo constante y fiable en los mercados, un desafío logístico y técnico, especialmente, si se considera la posible presencia de OVM en productos importados o procesados que pueden entrar en contacto con granos nativos (Vergaray et al., 2023).
Precisamente, para la detección de OVM en campo o mercados, se suelen emplear métodos de tamizaje rápido, como las tiras reactivas de flujo lateral (un tipo de prueba inmunocromatografíca), que detectan proteínas transgénicas específicas (Envirologix & Biotecnómica, 2019) y, si bien son herramientas económicas e inmediatas, su fiabilidad en matrices complejas son todavía una preocupación pues existe el riesgo de obtener falsos positivos debido a probables reacciones cruzadas no específicas o por la detección de contaminación superficial (polvo de grano transgénico), en lugar de una modificación genética real en el grano nativo analizado (Zhang et al., 2023), o escape de transgenes de cultivos hospedantes a parientes silvestres o variedades locales (Stewart, 2005).
El objetivo de este estudio fue detectar la presencia de OVM en maíz “chullpi” y “morado” de mercados de Lima Sur, evaluando críticamente la concordancia entre los resultados de las tiras de flujo lateral (Quick Comb®) y la confirmación molecular por rt-PCR. La relevancia de este estudio radica en la revelación de la inconsistencia de la reciprocidad de los resultados positivos de los métodos de tamizaje de las proteínas con la no detección de las secuencias transgénicas de esas proteínas en los granos de maíz evaluados, lo que podría ser una evidencia práctica que permita replantear los métodos de monitoreo que brinden mayor fiabilidad en los resultados, subrayando la necesidad de usar métodos moleculares para la vigilancia de la bioseguridad en el Perú.
La literatura proporciona información acerca de novedosos procedimientos para la detección de proteínas transgénicas en cultivos de maíz, aunque la especificidad de su determinación reduce el espectro de detección. Así se han ensayado inmunosensores para detectar proteínas Cry1Ab que inmoviliza anticuerpos policlonales anti-Cry1Ab de manera covalente midiéndose mediante cuantificación del voltaje cíclico y espectroscopia de impedancia electroquímica obteniéndose resultados muy precisos en la detección proteica (Freitas et al., 2016) , lo que resulta prometedor en contextos europeos, en donde también se proponen biosensores con nanotecnología para la detección de transcriptomas de plantas (Kumar et al., 2010; Kumar & Arora, 2020). No obstante, ambas tecnologías aún son muy costosas para nuestro país.
Por otra parte, en la búsqueda de métodos de detección más económicos y efectivos, se encontró un ensayo de detección amplia de las familias de proteínas Cry1 y Cry2 basado en el modelo de sándwich receptor-anticuerpo empleando HaCad-TBR como moléculas de captura y anticuerpos monoclonales como anticuerpos detectores. Estos métodos se validaron en cultivos de arroz y maíz en donde también dieron resultados positivos (Shen et al., 2024) siendo mucho más económico para su aplicación en campo. Coincidiendo con métodos de imagen espectral y machine learning en semillas de arroz que contenían los genes Cry1Ab y Cry1Ac que resultó ser un método económico y sobre todo efectivo en la detección de estos genes (Zhang et al., 2023)
Los estudios indican que no siempre hay coheren-cia entre los resultados de detección de proteínas transgénicas con la información detectada de las secuencias genómicas que las expresan. Ante esto existe la probabilidad de que la cantidad o número de copias de las secuencia transgénicas no sean suficientes para la amplificación y posterior secuenciamiento, por lo que podría optarse por el empleo de otras metodologías como la espectro-metría cercana al infrarrojo (Cicero Ribeiro et al., 2024). Sin embargo, señalan la imperiosidad de la realización de exámenes de eficacia de la detección, estableciendo estrategias de identificación y confirmación en contraste con los falsos positivos y la desaparición de los genes marcadores (Ahmed, 2002).
METODOLOGÍA
Diseño del estudio y recolección de muestras
Se realizó un estudio cuantitativo, con diseño no experimental de corte transversal, entre mayo y julio de 2024. Se seleccionaron 30 mercados de abastos mediante un muestreo intencional en seis distritos de Lima Sur, Perú: Lurín, Villa María del Triunfo (VMT), San Juan de Miraflores (SJM), Santiago de Surco, Villa el Salvador (VES) y Chorrillos. La selección se basó en el Censo Nacional de Mercados de Abastos 2016 (INEI, 2017) y criterios de inclusión como ser mercados mixtos o minoristas de alta afluencia. Se colectaron un total de 60 muestras (Aprox. 1 kg por muestra), correspondiendo a 30 muestras de maíz “chullpi” (Zea mays L.) y 30 de maíz “morado” (Zea mays L.), adquiridas a granel directamente de los puestos de venta.
Prueba de tamizaje (Inmunocromatografía)
Se utilizó el kit inmunocromatográfico QuickComb® (Envirologix Inc., EE. UU.) para la detección cualitativa de nueve proteínas transgénicas: Cry1Ab, CP4EPSPS, Cry3Bb, Cry1F, PAT/bar, Cry34, mCry3A, Cry2A y Vip3A. El análisis se realizó en el laboratorio de evaluación ambiental y diversidad biológica (EVADIB) de la Universidad Nacional Tecnológica de Lima Sur (UNTELS) siguiendo el protocolo del fabricante (Biotecnómica & Envirologix 2019).
Para cada una de las 60 muestras, se preparó un extracto moliendo 200 g de grano (Molino Marca Oster, Modelo BVSTBMH23-053) y añadiendo agua bidestilada en una proporción de 1:1.5 (peso/volumen). Debido a la diferencia en la textura del grano, el maíz “chullpi” se procesó con un molino (Molino Marca Oster, Modelo BVSTBMH23-053) y el maíz “morado” con una licuadora (Oster®, Modelo ActiveSense – 2081365 / BLSTTDGNBG) por un intervalo de 2 minutos. Las tiras reactivas se sumergieron en el extracto y los resultados se interpretaron visualmente tras 5 minutos.
Prueba de confirmación (PCR en tiempo real)
Las 60 muestras fueron agrupadas en siete conglomerados (5 de maíz “chullpi” y 2 de maíz “morado”), definidos por el tipo de maíz y el resultado (positivo/negativo) obtenido en la prueba de tamizaje. Estos conglomerados fueron analizados en el Laboratorio de Detección de OVM del Instituto Nacional de Innovación Agraria (INIA), acreditado bajo la norma NTP-ISO/IEC 17025. Se realizó la extracción de ADN y la detección de secuencias transgénicas mediante PCR en tiempo real (rt-PCR). La prueba se centró en la detección de los elementos reguladores más comunes en OVM comerciales: el promotor P-35S (del Virus del Mosaico de la Coliflor) y el terminador T-NOS (de Agrobacterium tumefaciens). El análisis incluyó los controles de calidad respectivos (control positivo, control negativo y control de extracción) para validar cada corrida.
Análisis de datos
Todos los análisis estadísticos y la generación de figuras se realizaron en el entorno de programación R v.4.5.1. Para evaluar la asociación entre el tipo de maíz (Chullpi/Morado) y la frecuencia de detección de cada proteína en el tamizaje, se utilizó la Prueba Exacta de Fisher, método preferido sobre Chi-cuadrado para tablas de contingencia con conteos bajos o nulos. Se aplicó un Análisis de Clúster Jerárquico (método de enlace "complete", distancia "binary") para visualizar el agrupamiento de las proteínas detectadas. Finalmente, se realizó una Inferencia Bayesiana Binomial, utilizando un prior no informativo (Beta(1,1)), para estimar la distribución posterior de la tasa de incidencia de los resultados positivos
RESULTADOS Y DISCUSIÓN
Detección por tamizaje
(Inmunocromatografía)
El tamizaje preliminar de las 60 muestras mediante tiras de flujo lateral (QuickComb®) arrojó una alta frecuencia de resultados positivos. En total, 11 de 30 (36,7%) muestras de maíz “chullpi” y 15 de 30 (50%) muestras de maíz “morado” reaccionaron positivamente a una o más de las nueve proteínas transgénicas evaluadas.
La distribución de las proteínas detectadas varió significativamente entre los dos tipos de maíz (Tabla 1).
Tabla 1
Frecuencia de detección (%) de proteínas transgénicas en muestras de maíz mediante inmunocromatografía
|
Proteína detectada |
Porcentaje de Positivo |
|
|
Maíz Chullpi |
Maíz Morado |
|
|
Cry1Ab |
30,0 |
36,7 |
|
CP4EPSPS |
6,7 |
10,0 |
|
Cry3Bb |
13,3 |
0,0 |
|
Cry1F |
36,7 |
16,7 |
|
PAT/bar |
36,7 |
0,0 |
|
Cry34 |
0,0 |
0,0 |
|
mCry3A |
26,7 |
6,7 |
|
Cry2A |
3,3 |
0,0 |
|
Vip3A |
3,3 |
50,0 |
En el maíz “chullpi”, las proteínas con mayor incidencia fueron PAT/bar (n=11, 36,7%) y Cry1F (n=11, 36,7%), seguidas de Cry1Ab (n=9, 30%) y mCry3A (n=8, 26,7%). En contraste, el maíz “morado” mostró una reacción dominante a la proteína Vip3A (n=15, 50%), seguida de Cry1Ab (n=11, 36,7%) (Figura 1). La proteína Cry34 no fue detectada en ninguna muestra.
Confirmación por PCR en Tiempo Real
En marcado contraste con los resultados del tamizaje, el análisis molecular confirmatorio arrojó resultados uniformemente negativos. Todas las 60 muestras, analizadas en 7 conglomerados (5 de Chullpi, 2 de Morado), resultaron negativas para la detección de las secuencias reguladoras P-35S y T-NOS (Tabla 2).
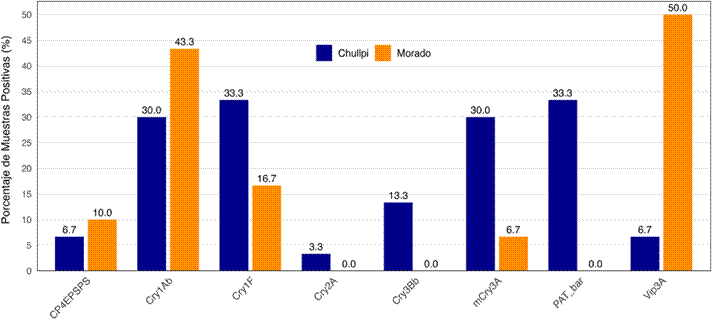
Figura 1. Porcentaje de muestras positivas (%) para cada proteína transgénica detectada por inmunocromatografía, comparando maíz Chullpi (n = 30) y maíz Morado (n = 30).
Tabla 2
Confirmación molecular (rt-PCR) para las secuencias P-35S y T-NOS en 7 conglomerados de muestras
|
N° Conglomerado |
Maíz |
Mercados |
Peso total (g) |
Resultado preliminar |
Resultado PCR |
|
|
1 |
Chullpi
|
● Jorge Chávez - Santiago de Surco ● ATECA - V.M.T ● Flores de Villa - S.J.M ● Cooperativa - V.M.T |
800 |
Positivo |
Negativo |
|
|
2 |
Chullpi |
● Cristo de Pachacamilla - V.E.S ● Santa Leonor - V. E.S. ● San Pedro- LURÍN ● Señor de Muruhuay - Santiago de Surco ● Trébol- Santiago de Surco ● San Pedro - Chorrillos |
800 |
Positivo |
Negativo |
|
|
3 |
Chullpi |
● Ciudad de Dios- S.J.M ● 1 de Abril - S.J.M ● Umamarca - S.J.M ● Modelo- S.J.M ● Amazonas - S.J.M ● San Roque - Santiago de Surco |
800 |
Negativo |
Negativo |
|
|
4 |
Chullpi |
● Villa Sur - V.E.S ● Unicachi - V.E.S ● Virgen de Cocharcas - V.E.S ● 5 de Junio - V.E.S ● San Martín - Chorrillos ● Kiwi - Chorrillos ● Mach - Chorrillos |
800 |
Negativo |
Negativo |
|
|
5 |
Chullpi |
● Las Palmas - V.M.T ● Plaza Micaela - V.M.T ● Virgen de Lourdes - V.M.T ● Santa Teresita - V.M.T ● Virgen de las Mercedes - Lurín ● Villa Alejandro - Lurín |
800 |
Negativo |
Negativo |
|
|
6 |
Morado |
● San Roque - Surco Amazonas - S.J.M ● 5 de Junio - V.E.S |
810 |
Negativo |
Negativo |
|
|
7 |
Morado |
● San Martín - Chorrillos ● Villa Alejandro - Lurín ● Santa Teresita - V.M.T |
800 |
Positivo |
Negativo |
Todos los ensayos de rt-PCR fueron validados por la correcta amplificación de los controles positivos internos, confirmando que la ausencia de detección en las muestras no se debió a inhibición de la reacción (Figura 2).
Análisis estadístico
Dado que la confirmación por PCR estableció que los granos no poseían las secuencias transgénicas, los resultados positivos de las tiras se analizaron como eventos de "falso positivo" para caracterizar su patrón.
1. Asociación de proteínas por tipo de maíz
El análisis de asociación (Prueba Exacta de Fisher) sobre los resultados del tamizaje (Tabla 3) reveló una fuerte correlación entre el tipo de maíz y el perfil de proteínas falsamente detectadas. La detección de PAT/bar (p < 0,001) y mCry3A (p = 0,042) estuvo significativamente asociada al maíz “chullpi”. La detección de Vip3A (p < 0,001) estuvo asociada casi exclusivamente al maíz “morado”. Las proteínas Cry1Ab, CP4EPSPS y Cry1F no mostra-ron una asociación estadísticamente significativa con un tipo de maíz específico.
2. Agrupamiento de proteínas detectadas (dendrogramas)
El análisis de clúster jerárquico visualiza cómo se agruparon las proteínas detectadas en las muestras positivas.
Maíz “chullpi” (Figura 3): Se observó una fuerte agrupación (baja distancia) entre las proteínas Cry3Bb y mCry3A, indicando que tendían a aparecer juntas. Las proteínas PAT/bar y Cry1F también formaron un clúster.
Maíz “morado” (Figura 4): Las proteínas detectadas formaron dos grupos principales. El primero incluyó a Cry1Ab y Vip3A (las más comunes), y el segundo a Cry1F, mCry3A y CP4EPSPS.
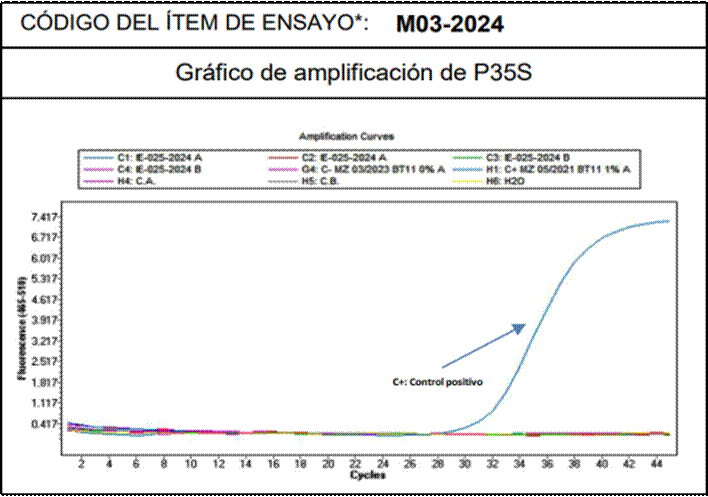
Figura 2. Gráfico de amplificación de rt-PCR representativo para la detección del promotor P-35S en el Conglomerado 1 (Chullpi). Se muestra la curva del control positivo (C+) y la ausencia de amplificación en la muestra (líneas planas), validando el resultado negativo.
Tabla 3
Resultados de la Prueba Exacta de Fisher para la asociación entre el tipo de maíz y la detección de proteínas (falsos positivos) mediante inmunocromatografía. Se muestra el valor p, el odds ratio estimado y el intervalo de confianza (IC) del 95%
|
Proteína |
p-value |
Odds ratio estimate |
Conf. low |
Conf. high |
|
Cry1Ab |
0,42197538 |
0,56592333 |
0,1679858 |
1,8358714 |
|
CP4EPSPS |
1 |
0,64756924 |
0,05040313 |
6,12000841 |
|
Cry3Bb |
0,11239964 |
- |
0,68915348 |
- |
|
Cry1F |
0,23259847 |
2,46187765 |
0,63979112 |
10,7574426 |
|
PAT_bar |
0,00079701 |
- |
2,88282373 |
- |
|
Cry34 |
1 |
0 |
0 |
- |
|
mCry3A |
0,04190012 |
5,83198722 |
1,05050672 |
61,0680573 |
|
Cry2A |
1 |
- |
0,02564066 |
- |
|
Vip3A |
0,00037167 |
0,07477878 |
0,00734211 |
0,38639959 |
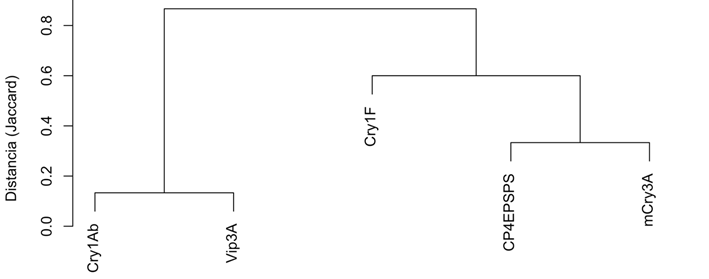
Figura 3. Dendrograma del Análisis de Clúster Jerárquico para las proteínas detectadas en maíz Morado (n=30). (Distancia: binary, Método: complete).

Figura 4. Dendrograma del Análisis de Clúster Jerárquico para las proteínas detectadas en maíz Chullpi (n=30). (Distancia: binary, Método: complete).
3. Inferencia de Incidencia Bayesiana
El análisis Bayesiano estimó la tasa de incidencia de los falsos positivos más significativos.
· Para Vip3A en maíz “morado” (k=15, n=30), la tasa de incidencia posterior media se estimó en 50,0%, con un intervalo creíble (IC) del 95% entre 33,5% y 66,5% (Figura 5A).
· Para PAT/bar en maíz “chullpi” (k=11, n=30), la media posterior fue de 36,7%, con un IC 95% entre 20,8% y 54,3% (Figura 5B).
· Para mCry3A en maíz “chullpi” (k=8, n=30), la media posterior fue de 26,7%, con un IC 95% entre 12,8% y 43,4% (Figura 5C).
El hallazgo principal de este estudio es la total discrepancia entre el tamizaje por inmunocro-matografía y la confirmación molecular. Mientras que las tiras rápidas (QuickComb®) arrojaron una alta tasa de detección (36.7% en maíz Chullpi y 50% en maíz Morado), el análisis confirmatorio por rt-PCR resultó 100% negativo para las secuencias transgénicas P-35S y T-NOS.
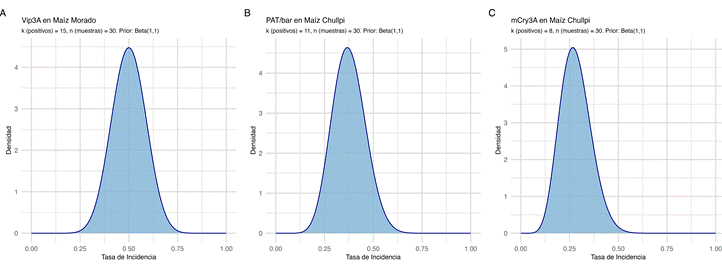
Figura 5. Distribuciones de probabilidad posterior (Inferencia Bayesiana) para la tasa de incidencia de falsos positivos en las proteínas más significativas. (A) Vip3A en maíz “morado” (k=15), (B) PAT/bar en maíz “chullpi” (k=11), y (C) mCry3A en maíz “chullpi” (k=8).
Esto lleva a asumir que, probablemente, todos los resultados positivos de las tiras sean falsos positivos. La discusión se centra en explicar las causas probables de esta discrepancia y sus profundas implicaciones para la bioseguridad en el Perú.
En el maíz Chullpi, los posibles falsos positivos se caracterizaron por un patrón de múltiples proteínas (Cry1Ab, Cry1F, PAT/bar, mCry3A, etc.). Sosteniendo que, muy probablemente estos resultados no se deban a un error del kit, sino a la detección de contaminación cruzada superficial, que ocurre desde el proceso de comercialización y transporte del maíz hasta los mercados de abastos. Las tiras QuickComb® son extremadamente sensibles y detectan la proteína transgénica, no el ADN del grano (Envirologix & Biotecnómica, 2019). La metodología de PCR, en cambio, incluyó el lavado externo de los granos antes de la extracción de ADN. Por lo tanto, es plausible que las tiras detectaran polvo o residuos de proteína transgénica adheridos a la superficie del grano nativo, mientras que el grano en sí mismo no era transgénico.
Esta hipótesis es consistente con el contexto de comercialización. Los granos se venden a granel en mercados donde el acopio, transporte y almacenamiento pueden facilitar la mezcla con polvo de maíz transgénico importado. Esta idea se ve reforzada por otros estudios, quienes, analizaron productos procesados e importados en Lima, confirmando mediante rt-PCR la presencia de OVM (Méndez & Vergaray, 2023). Nuestro estudio complementa ese hallazgo, sugiriendo que, aunque el grano nativo (Chullpi) está limpio, el riesgo de contaminación en la cadena de suministro es real y suficiente para generar resultados erróneos en pruebas de tamizaje sensibles. El patrón en el maíz “morado” fue notablemente diferente. Los posibles falsos positivos estuvieron dominados casi exclusivamente por la proteína Vip3A (50% de las muestras) y, en menor medida, Cry1Ab (36.7%). El análisis estadístico (Prueba de Fisher, Tabla 3) confirmó una asociación extremadamente fuerte entre el maíz “morado” y la detección de Vip3A.
Se propone que esto se deba a una interferencia de matriz no específica. El maíz “morado” se caracteriza por su alta concentración de antocianinas, el pigmento que le da su color característico (Bhushan et al., 2024). Es altamente probable que estos compuestos polifenólicos, o una alta concentración de proteínas endógenas en esta variedad, interfieran con los anticuerpos monoclonales de la tira reactiva de Vip3A, provocando una reacción cruzada o una "línea fantasma" que se interpreta como positiva. Este es un hallazgo metodológico crítico, ya que sugiere que el kit QuickComb®, aunque validado para maíz amarillo industrial, no sería fiable para variedades nativas altamente pigmentadas sin una validación específica. Esta hipótesis de interferencia por antocianinas, aunque fuertemente sugerida por los datos, requeriría una confirmación bioquímica específica.
Aunque los resultados de PCR fueron concluyentes, este estudio tiene limitaciones. Primero, el muestreo se restringió a mercados de Lima Sur; se requieren estudios en zonas de cultivo y otros mercados para una representatividad nacional. Segundo, el análisis de rt-PCR se centró en las secuencias P-35S y T-NOS; aunque cubren la gran mayoría de OVM comerciales, no se puede descartar la existencia teórica de eventos transgénicos con otros elementos reguladores, pues los OVM generalmente controlan sus transgenes a través de promotores constitutivos muy fuertes, pero que, en la expresión genética se ven influenciados por una gran cantidad de factores que aún no se han estudiado detalladamente en cultivos de maíz (Bakó et al., 2013).
Por otra parte, el potencial de detección proteico de los inmunoensayos muestra fortalezas importantes por la gran cobertura de proteínas detectables y por la capacidad de contraste con métodos más precisos a nivel de laboratorio. No obstante para obtenerse una mayor precisión en la detección es importante disponer de proteínas patentadas que actúen como estándares durante el desarrollo de las pruebas, lo que podrá garantizar un rendimiento uniforme en las pruebas implementadas en campo, validándose la variabilidad de la sensibilidad de las pruebas en diversos cultivos (Stave, 1999), condiciones que en el Perú aún no se implementan.
Asimismo, la recomendación de los monitoreos de proteínas expresadas en cultivos no solo deberían centrarse en algunas reconocidas sino que debería ampliarse a análisis proteómicos porque quepa la probabilidad de la expresión de rutas bioquímicas potencialmente dañadas que contribuirían en la diferenciación fenotípica proteica (Mercer & Wainwright, 2008; Varunjikar et al., 2023), pudiéndose detectar tempranamente y minimizar el impacto en los cultivos nativos y reducir la segregación genética en éstos, así en n casi 30 años de comercialización de cultivos GM, no existe evidencia de que las proteínas introducidas hayan causado alergias alimentarias en humanos (Ladics & Silvanovich, 2026).
Asumiendo que los resultados de rt-PCR en granos nativos sean verdaderamente negativos sería, en primera instancia, una noticia positiva. Sugiriendo que, a nivel de grano comercializado en Lima Sur, la Ley Nº 31111 estaría siendo efectiva y no se detecta un flujo genético de OVM hacia estas variedades. Sin embargo, la alta tasa de probables falsos positivos de las tiras de tamizaje genera una seria advertencia para los programas de vigilancia y bioseguridad. Un monitoreo basado únicamente en estas tiras habría generado una lamentable falsa alarma, declarando erróneamente una contaminación masiva de OVM en variedades nativas. Esto subraya la absoluta necesidad de que todo resultado positivo de un tamizaje inmunocromatográfico sea confirmado mediante métodos moleculares (rt-PCR), tal como lo realizan laboratorios acreditados (Singh et al., 2025).
No obstante, este desafío no es exclusivo de Perú, pues para el caso de Bolivia, la implementación de normativas de bioseguridad (como el Protocolo de Cartagena) requiere no solo un marco legal, sino también herramientas de diagnóstico validadas y fiables (Cartagena, 2020). Nuestros hallazgos demuestran que la aplicación directa de herramientas de diagnóstico importadas a nuestra biodiversidad nativa (maíces pigmentados) pueden fallar, y refuerza la necesidad de establecer protocolos de monitoreo adaptados a nuestra realidad.
CONCLUSIONES
El análisis de 60 muestras de maíz “chullpi” y “morado” de mercados de Lima Sur confirmó la ausencia de OVM en los granos, ya que no se detectaron secuencias transgénicas reguladoras (Promotor-35S y Terminador-NOS) mediante rt-PCR. Este resultado contrasta marcadamente con el tamizaje por inmunocromatografía (tiras QuickComb®), el cual mostró una alta frecuencia de detección (36,7% en Chullpi, 50% en Morado) que se concluyen como falsos positivos.
El patrón de estos falsos positivos difirió significativamente por variedad: en maíz chullpi”, la detección de múltiples proteínas sugiere una contaminación cruzada superficial con polvo de grano transgénico; mientras que, en maíz “morado”, la fuerte asociación con la proteína Vip3A indicaría una probable interferencia de matriz causada por antocianinas.
Este estudio demostró que los métodos de tamizaje rápido por inmunocromatografía no son fiables para la vigilancia de OVM en maíces nativos pigmentados o comercializados a granel. Se concluye que la confirmación molecular mediante rt-PCR es indispensable para la vigilancia de la bioseguridad en el Perú, a fin de evitar diagnósticos erróneos.
AGRADECIMIENTOS
Esta investigación fue financiada por la Univer-sidad Nacional Tecnológica de Lima Sur – RR N° 105-2023-UNTELS-R – Segunda Convocatoria para Proyectos de Investigación 2023.
REFERENCIAS BIBLIOGRÁFICAS
Ahmed, F. E. (2002). Detection of genetically modified organisms in foods. Trends in Biotechnology, 20(5), 215–223. https://doi.org/10.1016/S0167-7799(01)01920-5
Bakó, A., Gell, G., Zámbó, Á., Spitkó, T., Pók, I., Pintér, J., & Balázs, E. (2013). Monitoring transgene expression levels in different genotypes of field grown maize (Zea mays L.). South African Journal of Botany, 84, 6–10. https://doi.org/10.1016/J.SAJB.2012.09.005
Bhushan, B., Kumar, S., Kaur, C., Devi, V., Chaudhary, D. P., Singh, A., Dagla, M. C., Karjagi, C. G., Saleena, L. A. K., Chandran, D., & Kumar, M. (2024). Beyond colors: The health benefits of maize anthocyanins. Applied Food Research, 4(1), 100399. https://doi.org/10.1016/J.AFRES.2024.100399
Cartagena, P. (2020). Producción de transgénicos en Bolivia: Expectativas y problemas. Mundos Rurales, 15(1), 49–64. https://cipca.org.bo/docs/publications/es/237_ok-produccion-de-transgenicos-en-bolivia-expectativas-y-problemas.pdf
Cicero Ribeiro, M., Cabral, J., Nicolodelli, G., Senesi, G. S., Caires, A. R. L., Gonçalves, D. A., Menegatti, C., Milori, D., Cena, C., & Marangoni, B. (2024). Discrimination of maize transgenic and non-transgenic varieties by laser induced spectroscopy (LIBS) and machine learning algorithms. Microchemical Journal, 203, 110898. https://doi.org/10.1016/J.MICROC.2024.110898
Congreso de la República del Perú. (2010). Ley de Código de Protección y Defensa del Consumidor (p. 17). Congreso de la República del Perú. https://cdn.www.gob.pe/uploads/document/file/1351847/1244218-ley-29571_spij.pdf?v=1745616997
Congreso de la República del Perú. (2021). Ley No 31111 Modificatoria de la Ley 29811, Ley que establece la moratoria al ingreso y producción de organismos vivos al territorio nacional por un período de 15 años, a fin de establecer la moratoria hasta el 31 de diciembre de 2035 (p. 12). Presidencia del Consejo de Ministros. https://busquedas.elperuano.pe/dispositivo/NL/1917468-1
Envirologix, & Biotecnómica. (2019). Ficha Técnica Kit Quick CombTM para maíz en grano a granel.
Freitas, M., Correr, W., Cancino-Bernardi, J., Barroso, M. F., Delerue-Matos, C., & Zucolotto, V. (2016). Impedimetric immunosensors for the detection of Cry1Ab protein from genetically modified maize seeds. Sensors and Actuators B: Chemical, 237, 702–709. https://doi.org/10.1016/J.SNB.2016.06.149
INEI. (2017). Censo Nacional de Mercado de Abastos 2016 (p. 184). Instittuo Nacional de Estadística e Informática. https://www.inei.gob.pe/media/MenuRecursivo/publicaciones_digitales/Est/Lib1448/libro.pdf
Kumar, R., Singh, C. K., Kamle, S., Sinha, R. P., Bhatnagar, R. K., & Kachru, D. N. (2010). Development of nanocolloidal gold based immunochromatographic assay for rapid detection of transgenic vegetative insecticidal protein in genetically modified crops. Food Chemistry, 122(4), 1298–1303. https://doi.org/10.1016/J.FOODCHEM.2010.03.086
Kumar, V., & Arora, K. (2020). Trends in nano-inspired biosensors for plants. Materials Science for Energy Technologies, 3, 255–273. https://doi.org/10.1016/J.MSET.2019.10.004
Ladics, G. S., & Silvanovich, A. (2026). Protein Allergy and Genetically Engineered Crops. Reference Module in Biomedical Sciences, 199–234. https://doi.org/10.1016/B978-0-323-95488-4.00049-8
Méndez, C. R., & Vergaray, G. (2023). Detección y Cuantificación del Promotor 35S en productos alimenticios de maíz mediante PCR en tiempo real en Lima, Perú. Cátedra Villarreal Posgrado, 2(1), 39–51. https://doi.org/10.24039/RCVP2023211676
Mercer, K. L., & Wainwright, J. D. (2008). Gene flow from transgenic maize to landraces in Mexico: An analysis. Agriculture, Ecosystems & Environment, 123(1–3), 109–115. https://doi.org/10.1016/J.AGEE.2007.05.007
MINAM. (2018). Línea de Base de la diversidad genética del maíz peruano con fines de bioseguridad. Ministerio del ambiente. https://bioseguridad.minam.gob.pe/wp-content/uploads/2019/01/Linea-de-base-maíz-LowRes.pdf
Naqvi, S., Ramessar, K., Farré, G., Sabalza, M., Miralpeix, B., Twyman, R. M., Capell, T., Zhu, C., & Christou, P. (2011). High-value products from transgenic maize. Biotechnology Advances, 29(1), 40–53. https://doi.org/10.1016/J.BIOTECHADV.2010.08.009
Salvador-Reyes, R., & Clerici, M. T. P. S. (2020). Peruvian Andean maize: General characteristics, nutritional properties, bioactive compounds, and culinary uses. Food Research International, 130, 108934. https://doi.org/10.1016/J.FOODRES.2019.108934
Shen, C., Meng, M., Jin, J., Hu, X., Lin, M., Gao, M., Zhang, X., Zhu, Q., Xu, C., Xie, Y., Chen, C., Zhang, Z., Che, G., Pooe, O. J., Liu, Y., & Liu, X. (2024). Establishment of novel receptor-antibody sandwich assays to broadly detect Bacillus thuringiensis Cry1 and Cry2 toxins. International Journal of Biological Macromolecules, 254, 128034. https://doi.org/10.1016/J.IJBIOMAC.2023.128034
Singh, M., Shiwani, Aminedi, R., & Kaur, K. (2025). Detection method for CRISPR-Cas9 edited tomato with single nucleotide deletion using multiplex probe-based real-time PCR and confirmation of non-transgenicity: A case study. Journal of Genetic Engineering and Biotechnology, 23(4), 100609. https://doi.org/10.1016/J.JGEB.2025.100609
Stave, J. W. (1999). Detection of new or modified proteins in novel foods derived from GMO – future needs. Food Control, 10(6), 367–374. https://doi.org/10.1016/S0956-7135(99)00077-8
Stewart, C. N. (2005). Monitoring the presence and expression of transgenes in living plants. Trends in Plant Science, 10(8), 390–396. https://doi.org/10.1016/J.TPLANTS.2005.06.003
Varunjikar, M. S., Bøhn, T., Sanden, M., Belghit, I., Pineda-Pampliega, J., Palmblad, M., Broll, H., Braeuning, A., & Rasinger, J. D. (2023). Proteomics analyses of herbicide-tolerant genetically modified, conventionally, and organically farmed soybean seeds. Food Control, 151, 109795. https://doi.org/10.1016/J.FOODCONT.2023.109795
Vergaray, G., Méndez, C. R., Miguel, J. M., Gamboa, R. A., & Béjar, V. R. (2023). Determinación de transgenicidad y verificación en el etiquetado de alimentos industrializados de maíz en centros de expendio de Lima metropolitana. Anales de La Facultad de Medicina, 84(3), 279–285. https://doi.org/10.15381/ANALES.V84I3.25207
Zhang, J., Feng, X., Jin, J., & Fang, H. (2023). Concise Cascade Methods for Transgenic Rice Seed Discrimination using Spectral Phenotyping. Plant Phenomics, 5, 0071. https://doi.org/10.34133/PLANTPHENOMICS.0071
Zheng, J., Wang, W., Wang, W., Cui, T., Chen, S., Xu, C., & Engel, B. (2024). FACE-ing climate change: Propagation of risks and opportunities for cropping systems in mid-high-latitude regions: A case study between U.S. and China corn belts. Agricultural Systems, 220, 104087. https://doi.org/10.1016/J.AGSY.2024.104087