 |
INTRODUCCIÓN
PCR-RFLP In silico del gen 18S rRNA como alternativa para la identificación de nematodos entomopatógenos
In silico PCR-RFLP of the 18S rRNA gene as an alternative for the identification of entomopathogenic nematodes
Archi Alejandro Ruiz-Polo1 ; Joel Michel Riojas-Gonzales2, 3; Pedro S. Castillo-Carrillo2;
César Augusto Mogollón-Farias2, 3; Milton Valladolid-Ramos2; Segundo Melecio Garcia-Garcia2, 3;
Rosa Esmelda Cornejo-Hidalgo2; Cesar Alejandro Vasquez-Garcia3; Jose Stalyn Cordova Campos1, 2
1 Estación Experimental Agraria Los Cedros. Dirección de Recursos Genéticos y Biotecnología. Instituto Nacional de Innovación Agraria (INIA), Panamericana Norte kilómetro 12, Tumbes, Perú.
2 Facultad de Ciencias Agrarias, Universidad Nacional de Tumbes. Ciudad Universitaria, Av. Universitaria S/N, Tumbes, Perú.
3 Estación Experimental Agraria Los Cedros. Instituto Nacional de Innovación Agraria (INIA), Tumbes. Perú.
* Autor corresponsal: archi.ruiz.polo.mail.work@gmail.com (A. A. Ruiz-Polo).
ORCID de los autores
A. A. Ruiz-Polo: https://orcid.org/0009-0005-1273-2625 S. M. Garcia-Garcia: https://orcid.org/0009-0003-6300-9221
J. M. Michel Riojas-Gonzales: https://orcid.org/0009-0007-9966-9554 R. E. Cornejo-Hidalgo: https://orcid.org/0000-0002-2010-0442
P. S. Castillo-Carrillo: https://orcid.org/0000-0002-0255-1047 C. A. Vasquez-García: https://orcid.org/0009-0005-2803-7264
C. A. Mogollón-Farias: https://orcid.org/0009-0000-5390-9208 J. S. Cordova-Campos: https://orcid.org/0000-0002-5891-4679
M. Valladolid-Ramos: https://orcid.org/0000-0002-0526-0544
_______________________________________________________________________________________________________________________________________________
RESUMEN
En países en vías de desarrollo, el acceso a tecnologías modernas para el control de plagas es restringido, lo que plantea la necesidad de innovar en métodos y/o técnicas alternativas que sean más accesibles. El objetivo del estudio fue determinar la PCR-RFLP in silico del gen 18S ARNr como técnica alternativa en la identificación de nemátodos entomopatógenos. Se evaluaron dos cepas purificadas de larvas adultas de nematodos entomopatógenos, a partir de las cuales se extrajo el ADN genómico. Posteriormente, se llevó a cabo la amplificación del gen 18S del ARNr mediante PCR convencional y una secuenciación de ADN por la tecnología de Sanger en doble cadena. Las secuencias obtenidas fueron alineadas con el software MEGA v.11 y se generaron secuencias consenso de aproximadamente 850 pares de bases. Luego, se utilizaron herramientas como BLAST para la asignación taxonómica de especies y Restriction Mapper v.3 para el análisis de sitios de restricción y simulación de digestión enzimática. Las cepas analizadas se identificaron como Heterorhabditis indica y Heterorhabditis bacteriophora. En Heterorhabditis indica se hallaron 26 sitios de restricción, seleccionando cuatro sitios según su posición media (MslI, NruI, Tsp45I y BseRI), que luego de digerirlos in silico, generaron fragmentos de ADN con longitudes distintas. Por otra parte, en Heterorhabditis bacteriophora, se halló el mismo número de sitios de restricción, se seleccionaron tres (BglII, FokI y BstXI) y al digerirlos se obtuvieron fragmentos de ADN con diferentes longitudes. Los fragmentos de ADN (RFLP) obtenidos permitieron diferenciar claramente ambas especies. Los resultados demuestran que la técnica PCR-RFLP In silico del gen 18S ARNr es una herramienta efectiva para la identificación taxonómica de nematodos entomopatógenos, ofreciendo una alternativa viable en contextos donde los recursos son limitados y el control biológico es una alternativa.
Palabras clave: Entomología; Material genético; Plaga; PCR; RFLP.
In developing countries, access to modern pest control technologies is limited, highlighting the need to innovate in alternative methods and/or techniques that are more accessible. The aim of this study was to determine the in silico PCR-RFLP of the 18S rRNA gene as an alternative technique for the identification of entomopathogenic nematodes. Two purified strains from adult larvae of entomopathogenic nematodes were evaluated, from which genomic DNA was extracted. Subsequently, the 18S rRNA gene was amplified using conventional PCR, followed by double-stranded DNA sequencing via Sanger technology. The obtained sequences were aligned using MEGA v.11 software, and consensus sequences of approximately 850 base pairs were generated. Tools such as BLAST were then used for taxonomic species assignment, and Restriction Mapper v.3 was used for restriction site analysis and in silico enzymatic digestion simulation. The analyzed strains were identified as Heterorhabditis indica and Heterorhabditis bacteriophora. In Heterorhabditis indica, 26 restriction sites were found, with four selected based on their average position (MslI, NruI, Tsp45I, and BseRI), which, after in silico digestion, generated DNA fragments of different lengths. On the other hand, in Heterorhabditis bacteriophora, the same number of restriction sites was found, with three selected (BglII, FokI, and BstXI), and digestion also produced DNA fragments of varying lengths. The obtained DNA fragments (RFLPs) allowed for clear differentiation between the two species. The results demonstrate that the In silico PCR-RFLP technique of the 18S rRNA gene is an effective tool for the taxonomic identification of entomopathogenic nematodes, offering a viable alternative in contexts where resources are limited and biological control is a viable option.
Keywords: Entomology; Genetic material; Pest; PCR; RFLP.
Recibido: 05-08-2025.
Aceptado: 27-03-2026.
|
INTRODUCCIÓN
Los nematodos entomopatógenos han adquirido una notable relevancia en el ámbito del control biológico de plagas, debido a su capacidad de actuar como agentes letales contra insectos que afectan negativamente la productividad agrícola (Půža & Tarasco, 2023). Estos organismos establecen asociaciones simbióticas con bacterias de los géneros Xenorhabdus y Photorhabdus, las cuales son liberadas en el interior del hospedero tras la infección, provocando septicemia y la muerte del insecto en un corto periodo de tiempo (Gaugler, 2002). Esta característica los posiciona como herramientas prometedoras dentro de programas de manejo integrado de plagas, especialmente en sistemas agrícolas sostenibles (Koppenhöfer et al., 2020; Nurashikin-Khairuddin, et al., 2022).
Entre los principales nematodos entomopatógenos destacan especies de los géneros Heterorhabditis y Steinernema, ampliamente estudiadas por su eficacia biocontroladora (Yangüéz et al., 2024). Dentro de estas, Heterorhabditis indica (Shinde et al., 2022) y Heterorhabditis bacteriophora (Sánchez et al., 2019; Leonar et al., 2022) han demostrado alta virulencia frente a diversos insectos plaga. Asimismo, especies como Steinernema riobrave y Rhabditis blumi han sido reportadas como parásitos obligados, cuya fase infectiva juvenil penetra al hospedero a través de aberturas naturales, facilitando la colonización interna y posterior reproducción en el cadáver del insecto (Poinar, 1979; Gaugler, 2002).
La correcta identificación de estas especies resulta crucial, ya que su eficacia en el control biológico depende de factores como la especificidad del hospedero, condiciones ambientales y características genéticas propias de cada cepa (Rusinque et al., 2023; Rangarajan et al., 2025). Tradicionalmente, la identificación de nematodos se ha basado en características morfológicas; sin embargo, estas presentan limitaciones debido a la plasticidad fenotípica y similitudes entre especies (Bogale et al., 2020; Bhat et al., 2022). En respuesta a estas limitaciones, se han desarrollado enfoques moleculares basados en la amplificación y secuenciación de marcadores genéticos, como el gen 18S ARNr, ITS y COI, los cuales permiten una identificación más precisa (Valderrama-Aravena, 2025).
No obstante, técnicas como la secuenciación de ADN y la espectrometría de masas, aunque altamente precisas, implican elevados costos, requerimientos tecnológicos especializados y limitada accesibilidad en países en vías de desarrollo (Córdova-Campos et al., 2023). Esta situación restringe su aplicación en contextos donde el control biológico podría tener mayor impacto, generando la necesidad de alternativas metodológicas más accesibles, sin comprometer la confiabilidad en la identificación taxonómica.
En este contexto, la técnica de PCR-RFLP (Reacción en Cadena de la Polimerasa seguida de Polimorfismo de Longitud de Fragmentos de Restricción) se presenta como una herramienta eficiente, rápida y de bajo costo para la diferenciación de especies mediante el análisis de patrones de restricción del ADN (Chena et al., 2014; Torchia et al., 2021; Gao et al., 2023). Diversos estudios han demostrado su aplicabilidad en la identificación de organismos en distintos grupos biológicos, incluyendo nematodos, destacando su utilidad en entornos con recursos limitados (Peraza-Padilla et al., 2013; Montero et al., 2007; Madikadike et al., 2024). Asimismo, investigaciones recientes han señalado que, la PCR-RFLP posee un potencial alto para discriminar especies cercanas mediante el análisis de regiones conservadas en marcadores ribosomales (Niloufar, 2020).
A pesar de estos avances, existe un vacío en el conocimiento respecto a la validación sistemática de la técnica PCR-RFLP mediante enfoques in silico aplicados específicamente a nematodos entomopatógenos. En particular, son limitados los estudios que evalúan la capacidad discriminatoria de enzimas de restricción sobre secuencias del gen 18S ARNr en diferentes especies, lo que dificulta establecer protocolos estandarizados y reproducibles para su implementación en programas de control biológico.
En ese sentido, estudios bioinformáticos orientados a evaluar la viabilidad de la PCR-RFLP in silico resultan fundamentales, ya que permiten predecir patrones de restricción, optimizar la selección de enzimas y reducir costos experimentales. Además, su aplicación contribuiría al fortalecimiento de estrategias de manejo agroecológico, mediante la caracterización precisa y oportuna de nematodos entomopatógenos tanto autóctonos como introducidos.
Por lo tanto, el objetivo del presente estudio fue determinar la viabilidad de la técnica PCR-RFLP in silico del gen 18S ARNr como alternativa para la identificación de nematodos entomopatógenos.
METODOLOGÍA
Diseño del estudio
Se desarrolló una investigación compuesta por una fase in vitro y otra in silico, bajo un diseño no experimental, con enfoque cualitativo y nivel descriptivo. El estudio se basó en el análisis de bioinformática de las secuencias del ADN del gen ribosomal 18S de dos especímenes de nematodos entomopatógenos aislados en el norte de Perú. El diseño de la investigación y su flujo de trabajo se observa en la Figura 1, la cual describe en la metodología cada uno de los componentes.
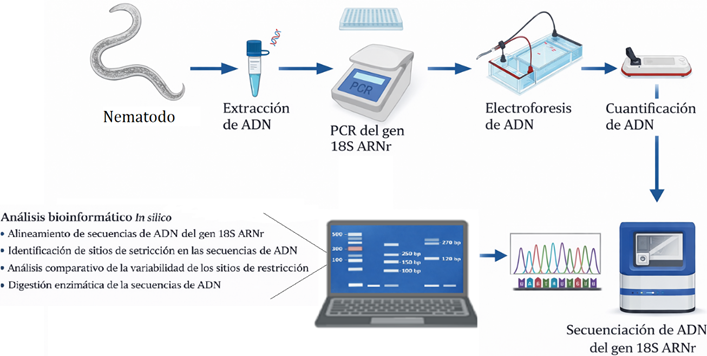
Figura 1. Esquema del diseño de la investigación In silico en la que se describe el flujo de trabajo desde el organismo vivo (nematodo) hasta la fragmentación del gen 18S ARNr mediante análisis bioinformáticos.
Material biológico
Dos cepas (H1 y H2) de larvas adultas de nematodos pertenecientes al género Heterorhabditis, purificadas y previamente enfrentadas al insecto plaga Galleria mellonella (polilla de cera), fueron proporcionadas por la Universidad Nacional de Tumbes. Las cepas corresponden a aislados de suelos asociados a Neltuma pallida, recolectadas en la región de Tumbes, Perú (3°34'1'' S, 80°27.092' O).
Extracción de ADN y PCR dirigida al gen 18S ARNr
Las cepas purificadas contenidas en una solución de suspensión, fueron sometidas a un protocolo de extracción de ADN genómico siguiendo las instrucciones del método estándar de un kit comercial. Luego, el gen 18S ARNr fue amplificado utilizando los cebadores SSU18A: AAAGATTAAGCCATGCATG y SSU26R: CATTCTTGGCAAATGCTTTCG. La reacción de PCR se preparó en un volumen total de 25 µL, compuesto por 15,4 µL de agua ultra pura, 2,5 µL de tampón 10X, 1,5 µL de MgCl₂ (25 mM), 0,5 µL de mezcla de dNTPs, 0,5 µL de cada cebador, 0,1 µL de Taq ADN polimerasa y 4 µL de ADN genómico.
El programa de amplificación consistió en una desnaturalización inicial a 95 °C durante 5 minutos, seguida de 35 ciclos con desnaturalización a 94 °C por 1 minuto, alineamiento a 52 °C por 1 minuto y 30 segundos, y extensión a 68 °C durante 2 minutos. Finalmente, se realizó una extensión final a 68 °C por 10 minutos y conservación a 4 °C por un máximo de 24 horas. Se incluyó un control negativo para descartar contaminación durante la amplificación, como producto PCR se esperó un amplicón con un tamaño de 850 pares de bases (pb).
Electroforesis en gel de agarosa
Los productos de PCR fueron evaluados mediante electroforesis en gel de agarosa al 1,5 %, preparado en solución tampón TAE 1X y teñido con bromuro de etidio a una concentración de 0,5 ng/mL. Para cada corrida se emplearon 8 µL del amplicón, 4 µL de tampón de carga y 2 µL de un marcador de peso molecular de 1000 pares de bases. La separación electroforética se realizó aplicando un voltaje de 70 V durante 30 min.
Secuenciación de ADN
Los productos amplificados por PCR fueron enviados a una empresa especializada en servicios biotecnológicos para su secuenciación mediante la tecnología Sanger de doble cadena. Las secuencias obtenidas fueron entregadas en formato FASTA, un archivo de texto plano estandarizado, utilizado para su posterior análisis de bioinformática. Posteriormente, las secuencias se analizaron mediante softwares de bioinformática.
Asignación taxonómica
Se realizó un alineamiento de secuencias de ADN utilizando el software Molecular Evolutionary Genetics Analysis (MEGA), versión 11. Las secuencias obtenidas de las lecturas directa (SSU18A) e inversa (SSU26R) fueron alineadas por complementariedad de bases, empleando el algoritmo MUSCLE.
Posteriormente, se depuraron los extremos no coincidentes, huecos y se seleccionó la región consenso, la cual fue exportada en formato FASTA a la herramienta básica de búsqueda de alineación local (BLAST, por sus siglas en inglés), con la que se realizó la asignación taxonómica de las secuencias de ADN a nivel de especie, mediante comparación de similitudes con secuencias disponibles en el banco de genes del Centro Nacional para la Información Biotecnológica (NCBI, por sus siglas en inglés). Los parámetros de homología considerados fueron una cobertura y similitud mínima del 95% y máxima del 100%. Finalmente, las secuencias fueron conservadas en formato FASTA para la identificación de sitios de restricción.
Mapeo de sitios de restricción
Se realizó un mapeo de sitios de restricción coincidentes con endonucleasas homónimas en las secuencias del presente estudio y en las secuencias con las que se halló similitud. El mapeo se realizó empleando el software Restriction Mapper, versión 3. Los parámetros considerados fueron: confor-mación lineal del ADN, ordenamiento de enzimas por frecuencia de corte, filtrado limitado a un máximo de un sitio de corte, longitud mínima del sitio de reconocimiento de cinco pares de bases, inclusión de todas las enzimas comerciales y consideración de extremos cohesivos y extremos romos (-5’-3’-). Como producto, se obtuvo un mapa detallado de sitios de restricción.
Digestión enzimática In silico
La digestión enzimática fue simulada en software Restriction Mapper, v. 3. Previamente, se seleccionó un sitio de restricción ubicado aproximadamente en la región media de la secuencia consenso del gen, con el objetivo de generar fragmentos de ADN de tamaño representativo y fácilmente observables en aplica-ciones in vitro. Se fragmentó la secuencia total del gen en los sitios de restricción seleccionados y se obtuvieron fragmentos de longitud polimórfica (RFLP) con distintos tamaños según la posición de los sitios.
Aspectos éticos
Se utilizaron especímenes de nematodos prove-nientes de una tesis de la Universidad Nacional de Tumbes, adoptando consideraciones éticas, como la gestión de permisos y la mención del origen del material biológico analizado.
RESULTADOS Y DISCUSIÓN
Asignación taxonómica
Los productos amplificados por PCR del gen 18S ARNr fueron secuenciados y alineados, lo que permitió la construcción de secuencias consenso de 850 pares de bases tanto para el nematodo H1 como para H2. Estas secuencias fueron posterior-mente analizadas mediante la herramienta BLAST, donde se compararon con secuencias homólogas disponibles en la base de datos del NCBI. El análisis de similitud reveló una correspondencia taxonó-mica difusa, ya que a H1 y H2 se asignaron especies que no corresponden al género Heterorhabditis, tal como estaban previamente identificadas mediante taxonomía clásica.
No obstante, entre las otras especies asignadas, se observaron especies del género Heterorhabditis, por lo que se seleccionaron aquellas que poseen una cobertura y similitud máxima del 99%. Se tamizaron las similitudes con especies distintas a Heterorhabditis. Se seleccionaron las especies Heterorhabditis indica para H1 y Heterorhabditis bacteriophora para H2 (Tabla 1). Los resultados de la asignación taxonómica son consistentes con lo reportado por Córdova-Campos et al. (2025), quienes, mediante el análisis molecular del gen 18S ARNr, lograron identificar cepas de nematodos aisladas de suelos forestales. En dicho estudio, estas cepas fueron evaluadas frente a larvas de Galleria mellonella, determinándose qué las especies con efecto nematicida fueron Hetero-rhabditis indica y Heterorhabditis bacteriophora. Por otro lado, diversos autores han destacado la alta sensibilidad del gen 18S ARNr como marcador molecular en la identificación precisa de nemato-dos (Floyd et al., 2002; Vera-Morales et al., 2024), también señalado en estudios realizados en Perú (Córdova-Campos et al., 2023),
De acuerdo con Vélez & Guzmán (2022), además de los genes 18S del ADN ribosómico (ADNr), otras regiones genéticas comúnmente utilizadas para la identificación de nematodos de importancia agrícola incluyen secuencias del ADN mitocondrial (mtDNA) y regiones del espaciador intergénico (IGS), lo que permite una caracterización molecular más completa y precisa. Por su parte Chaves-Velásquez et al. (2023) destacan que tanto el gen 18S del ADN ribosómico como la región interna transcrita que abarca ITS1-5.8S-ITS2 constituyen herramientas moleculares eficaces para la identificación específica de nematodos fitopará-sitos asociados a cultivos de interés agrícola como el plátano y el banano.
Tabla 1
Determinación taxonómica de nematodos a nivel especie mediante análisis de similitud de secuencias de ADN del gen 18S ARNr
|
Cepa |
Especie |
% Cobertura |
% Identidad |
Nº Accesión |
|
H1 |
Heterorhabditis indica |
99 |
99,65 |
PQ608564.1 |
|
Heterorhabditis indica |
97 |
99,52 |
OL470132.1 |
|
|
Heterorhabditis indica |
97 |
99,28 |
OK562793.1 |
|
|
H2 |
Heterorhabditis bacteriophora |
95 |
100 |
JN996479.1 |
|
Heterorhabditis bacteriophora |
95 |
99,87 |
JN996475.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,54 |
AF036593.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,54 |
FJ040428.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,54 |
FJ040429.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,54 |
OR999415.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,42 |
FJ040430.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,42 |
KJ636408.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,42 |
KY299838.1 |
|
|
Heterorhabditis bacteriophora |
97 |
97,42 |
MK214461.1 |
En conjunto, estos hallazgos refuerzan la utilidad del gen 18S ARNr como herramienta confiable en estudios de sistemática molecular de nematodos entomopatógenos u otras especies como fitoparásitos.
Mapeo de sitios de restricción
Se identificaron diversos sitios de restricción a lo largo de la secuencia del gen 18S ARNr en las especies de nematodos analizadas. En Heterorhabditis indica se detectaron 30 sitios, de los cuales 26 se preseleccionaron, y posterior-mente se eligieron 3 (resaltados en amarillo) en función de su ubicación en posiciones cercanas a la región media de la longitud total de la secuencia, esto con el fin de que cuando la técnica se reproduzca In vitro, los RFLPs puedan ser visualizados como bandas con luminiscencia nítida en los geles de agarosa (Tabla 1).
De forma análoga, se identificaron 58 sitios de restricción en Heterorhabditis bacteriophora, de los cuales también se preseleccionaron 26, seleccio-nando finalmente 3 sitios (resaltados en verde) aplicando el mismo criterio de la posición media en la secuencia del gen (Tabla 2).
Según Inglehart & Nelson (1994), desde hace décadas los mapeos de sitios de restricción automatizados poseen limitaciones, puesto que al evaluarse 8 programas de mapeo, desarrollados hasta 1993, se evidenció la falta de distinción y fiabilidad de mapas verdaderos dado la manifes-tación de errores en la búsqueda de sitios de restricción. Sin embargo, estudios realizados durante los mismos años en Japón, han evidenciado que, la PCR-RFLP empleada con genes mitocon-driales puede ser eficiente, puesto que se ha permitido la identificación molecular precisa de las especies de nematodos M. hapla, M. suginamiensis, M. marylandi, M. mali, M. camelliae, M. sp. near mali y M. sp. near arenaria (Orui, 1998). Posteriormente, se demostró la existencia de mejoras sustanciales en la aplicación de esta técnica, por ejemplo, Solano et al. (2011), evidenciaron una alta precisión de sitios de restricción mapeados mediante el uso de enzimas específicas como Dra I y Hinf I, las cuales permitieron diferenciar especies de nematodos agalladores del género Meloidogyne asociados a plantas ornamentales, a partir de productos de PCR del gen mitocondrial Citocromo Oxidasa Subunidad I (COI). No obstante, actualmente en países como Estados Unidos, técnicas como la PCR convencional, aún sigue considerándose para la identificación del nemátodo agallador Meloidogyne spp, siendo el ADN mitocondrial y ribosomal las dianas moleculares empleadas (Ye et al., 2019; Ye et al., 2021).
Digestión enzimática in silico
La digestión enzimática in silico del gen 18S ARNr de Heterorhabditis indica, utilizando los sitios de restricción MslI, NruI, Tsp45I y BseRI, generó fragmentos de ADN de diferentes longitudes (Figura 2). De manera similar, la digestión computacional del gen 18S ARNr de Heterorhabditis bacteriophora con las endonucleasas de restricción BglII, FokI y BstXI produjo fragmentos de ADN con tamaños variables (Figura 3).
Tabla 2
Sitios de restricción en el gen ribosomal 18S de los nematodos entomopatógenos analizados
|
Nº |
Heterorhabditis indica |
|
Heterorhabditis bacteriophora |
||||
|
Enzima |
Secuencia |
Posición de corte |
|
Enzima |
Secuencia |
Posición de corte |
|
|
1 |
HpaI |
GTTAAC |
794 |
|
AccI |
GTMKAC |
267 |
|
2 |
MslI |
CAYNNNNRTG |
443 |
|
ApoI |
RAATTY |
821 |
|
3 |
NruI |
TCGCGA |
338 |
|
AvaII |
GGWCC |
829 |
|
4 |
PsiI |
TTATAA |
33 |
|
BccI |
CCATC |
245 |
|
5 |
AccI |
GTMKAC |
178 |
|
BglII |
AGATCT |
443 |
|
6 |
AvrII |
CCTAGG |
615 |
|
FokI |
GGATG |
326 |
|
7 |
BccI |
CCATC |
250 |
|
NcoI |
CCATGG |
271 |
|
8 |
BclI |
TGATCA |
282 |
|
SfaNI |
GCATC |
118 |
|
9 |
BsmAI |
GTCTC |
408 |
|
SmlI |
CTYRAG |
223 |
|
10 |
FauI |
CCCGC |
654 |
|
TfiI |
GAWTC |
579 |
|
11 |
MluI |
ACGCGT |
287 |
|
VspI |
ATTAAT |
768 |
|
12 |
NcoI |
CCATGG |
276 |
|
XhoI |
CTCGAG |
223 |
|
13 |
SalI |
GTCGAC |
177 |
|
BciVI |
GTATCC |
812 |
|
14 |
Tsp45I |
GTSAC |
398 |
|
BseMII |
CTCAG |
577 |
|
15 |
Tth111I |
GACNNNGTC |
586 |
|
BsgI |
GTGCAG |
156 |
|
16 |
XbaI |
TCTAGA |
90 |
|
BsrDI |
GCAATG |
530 |
|
17 |
BseRI |
GAGGAG |
405 |
|
BstXI |
CCANNNNNTGG |
419 |
|
18 |
BsmI |
GAATGC |
812 |
|
EciI |
GGCGGA |
202 |
|
19 |
NspI |
RCATGY |
106 |
|
Eco57I |
CTGAAG |
630 |
|
20 |
SacII |
CCGCGG |
509 |
|
GsuI |
CTGGAG |
115 |
|
21 |
AflIII |
ACRYGT |
222, 287 |
|
HaeIV |
GAYNNNNNRTC |
468 |
|
22 |
ApoI |
RAATTY |
836 |
|
MboII |
GAAGA |
449 |
|
23 |
AvaII |
GGWCC |
588, 756 |
|
MmeI |
TCCRAC |
537 |
|
24 |
EcoP15I |
CAGCAG |
388, 530 |
|
NmeAII |
GCCGAG |
617 |
|
25 |
PleI |
GAGTC |
174, 305 |
|
NspI |
RCATGY |
109 |
|
26 |
StyI |
CCWWGG |
276, 615 |
|
SacII |
CCGCGG |
500 |
n/r: “no registra”.
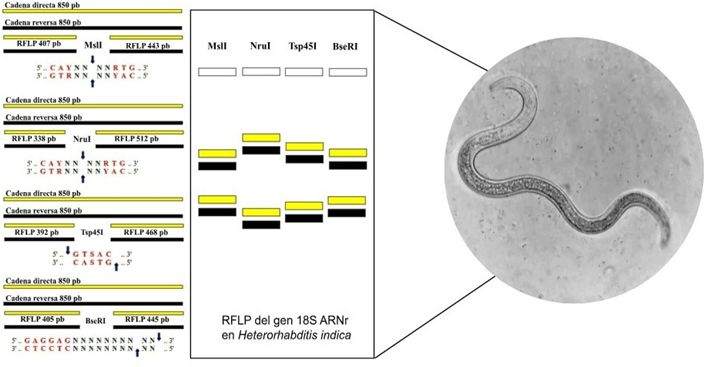
Figura 2. PCR-RFLP in silico del gen 18S ARNr de Heterorhabditis indica.
Los resultados demostraron qué, mediante los sitios de restricción seleccionados se obtienen RFLPs con tamaños suficientes como para ser visualizados en geles de agarosa in vitro, lo cual hace posible diferenciar ambas especies previamente asignadas por taxonomía molecular. Perteguer et al. (2004) aplicaron un enfoque similar para la caracterización de nematodos anisákidos provenientes de peces, utilizando regiones amplificadas del ITS-1, el gen del ARN ribosomal 5.8S, el ITS-2 y una porción del gen 28S, revelando que la digestión enzimática dirigida a sitios de restricción permitió no solo identificar la especie, sino también evidenciar la existencia de variabilidad genética intraespecífica. Otros autores como Rodríguez (2019), han identificado a Heterorhabditis bacteriophora y Heterorhabditis indica mediante la digestión enzimática del sitio de restricción Alu1 del gen 28S ARNr demostrando que dichas regiones de ADN permiten una identificación específica de nematodos.
A lo largo de los últimos años, se ha evidenciado un incremento sostenido en el uso de softwares especializados para el mapeo in silico de sitios de restricción, los cuales se han consolidado como herramientas fundamentales en diversas disciplinas (Bikandi et al., 2004; Laref et al., 2024). Estas plataformas permiten optimizar la precisión en la identificación y localización de sitios de corte a lo largo de secuencias genómicas completas (Ruiz-Polo, Luis Arismendiz et al., 2024; Chena et al., 2014), facilitando la generación de perfiles RFLP cuyas longitudes teóricas predichas muestran una alta concordancia con los patrones obtenidos experimentalmente en laboratorio (Wang et al., 2025; Ruiz-Polo, Santillán-Valdivia et al., 2024).
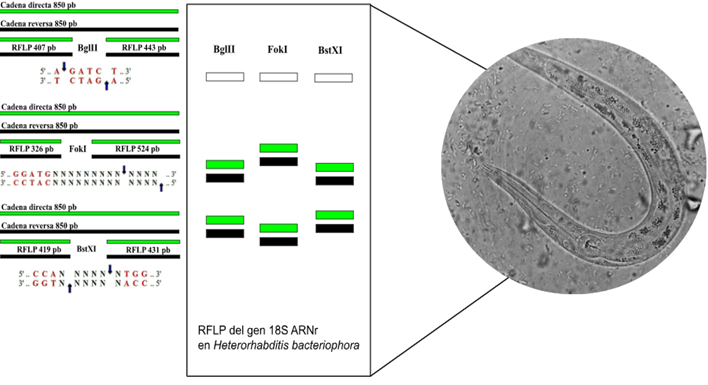
Figura 3. PCR-RFLP in silico del gen 18S ARNr de Heterorhabditis bacteriophora.
Tabla 3
Comparación entre las características de técnicas moleculares empleadas para la identificación de nematodos
|
Técnica molecular |
Características |
||||
|
Tiempo en la obtención de resultados |
Complejidad de uso |
Especificidad/ sensibilidad |
Fuente |
|
|
|
PCR convencional |
2 a 3 horas |
Requiere capacitación técnica especializada para el manejo de equipos y reactivos de extracción de ADN y amplificación de genes |
Sensibilidad: Moderada Especificidad: Moderada (depende de los primers) |
Floyd et al. (2005) Carta & Li (2018) Rodríguez (2019) Huang et al. (2023) |
|
|
qPCR |
2 a 4 horas |
Requiere capacitación técnica especializada para el manejo de equipos y reactivos de secuenciación |
Sensibilidad: Muy alta Especificidad: Muy alta |
Huang et al. (2007) Chen et al. (2022) Hodson et al. (2023) Huang et al. (2023) Goraya y Yan. (2024) Córdoba-Sánchez et al. (2026) |
|
|
PCR multiplex |
2 a 4 horas |
Requiere capacitación técnica especializada para el manejo de equipos y reactivos de secuenciación |
Sensibilidad: Moderada a alta Especificidad: Alta (si está bien optimizada) |
Fang et al. (2010) Hu et al. (2011) Huang et al. (2019) Ren et al. (2024) |
|
|
PCR convencional + Secuenciación de ADN |
2 a 10 días (dependiendo del país donde se secuencia) |
Requiere capacitación técnica especializada para el manejo de equipos y reactivos de secuenciación |
Sensibilidad: Alta Especificidad: Muy alta (Muy precisa) |
Según Lacey et al. (2001) Valderrama-Aravena et al. (2014) Chowdhury y Yan (2021) Córdova-Campos et al. (2023) Córdova-Campos et al. (2025) |
|
|
*PCR convencional – RFLP (Este trabajo) |
*2 a 4 horas |
*Requiere capacitación técnica especializada para el manejo de equipos y reactivos de extracción de ADN y amplificación de genes |
*Sensibilidad: Moderada *Especificidad: Moderada a alta *Menos precisa que la secuenciación (puede haber patrones similares). |
Perteguer (2004) Campos-Herrera, (2006) Susurluk et al. (2007) Montero et al. (2017) Gao et al. (2023) Ruiz-Polo, Santillán-Valdivia et al. (2024) Ruiz-Polo, Luis Arismendiz et al. (2024) Andree et al. (2024) |
|
(*) indica la técnica molecular In silico analizada en el presente estudio.
Sensibilidad: detectar verdaderos positivos.
Especificidad: excluir falsos positivos.
Andree et al. (2024) han reportado que los análisis basados en sitios de restricción, mediante PCR-RFLP, constituyen una estrategia eficiente para la diferenciación sexual en peces, ampliando así el espectro de aplicaciones de esta metodología en estudios de genética aplicada. Por tanto, considerando la bibliografía consultada, los resultados obtenidos en el presente estudio pueden ser dilucidados (Tabla 3). En cuanto a la digestión enzimática del gen 18S ARNr en el que se identificaron los sitios de restricción MslI, NruI, Tsp45I y BseRI para Heterorhabditis indica, y BglII, FokI y BstXI para Heterorhabditis bacteriophora, estudios previos han reportado que el sitio de restricción AluI se localiza en una región genética que permite discriminar entre nematodos entomopatógenos como Heterorhabditis bacteriophora y Heterorhabditis indica. No obstante, dicha identificación se ha realizado empleando como marcador molecular el gen 28S del ARN ribosomal (Rodríguez, 2019). Según Lacey et al. (2001), diversos métodos y marcadores moleculares han sido previamente empleados no solo con fines de diagnóstico de especies crípticas, poblaciones y cepas de nematodos, sino también para el análisis de sus relaciones filogenéticas y patrones evolutivos. Esto es respaldado por autores como Peteira et al. (2014), quienes señalan que, entre los métodos mayormente utilizados para identificar nematodos, se encuentra la reacción en cadena de la polimerasa (PCR) en sus dos variantes, la PCR-RFLP y la amplificación de ADN polimórfico al azar (RAPD), ya que puede ser usados de forma fácil por personas no expertas. Esto coincide con estudios de la última década en los que se ha trabajado como técnica molecular la PCR-RFLP para identificar especies de nemátodos entomopatógenos como Steinernema feltiae Heterorhabditis bacteriophora (Campos-Herrera, 2006; Susurluk et al., 2007).
CONCLUSIONES
Los resultados permitieron la identificación de sitios de restricción en una longitud de 850 pb del gen 18S ARNr en Heterorhabditis indica (MslI, NruI, Tsp45I, BseRI) y Heterorhabditis bacteriophora (BglII, FokI, BstXI), con los que se obtuvieron fragmentos de ADN (RFLP) que permiten su identificación molecular. Esto respalda su aplicación como una alternativa molecular eficaz y accesible para la diferenciación de especies de nemátodos, especialmente en contextos con limitaciones tecnológicas para la secuenciación de ADN, debido a su bajo costo y simplicidad operativa.
Por otro lado, el presente estudio presenta como limitación el análisis de longitudes polimórficas del gen 18S ARNr en nematodos nativos del departamento de Tumbes, por lo que los resultados obtenidos son directamente aplicables a poblaciones de Heterorhabditis indica y Heterorhabditis bacteriophora presentes en dicha región. Sin embargo, la metodología puede ser reproducida en otras zonas geográficas mediante la secuenciación del mismo gen y la evaluación de sus patrones de restricción, permitiendo validar su aplicabilidad en distintos contextos ecológicos.
En ese sentido, se espera que este estudio in silico contribuya como una herramienta alternativa para programas de vigilancia fitosanitaria en el Perú, facilitando la identificación rápida de nematodos de interés agrícola. Asimismo, se recomienda la aplicación de la técnica PCR-RFLP en áreas específicas donde aún existen vacíos de cono-cimiento, tales como: (i) la caracterización mole-cular de nematodos entomopatógenos nativos en agroecosistemas poco estudiados, (ii) la dife-renciación de especies crípticas o filogenética-mente cercanas que presentan ambigüedad morfológica, (iii) el monitoreo de la variabilidad genética de poblaciones de nematodos utilizadas en programas de control biológico, y (iv) la detección temprana de especies invasoras o introducidas en sistemas agrícolas. Estas aplica-ciones permitirían fortalecer el conocimiento taxonómico y ecológico de estos organismos, así como optimizar estrategias de manejo agroecológico sostenible.
AGRADECIMIENTOS
A la Universidad Nacional de Tumbes por la provisión de las cepas de nematodos utilizadas en el presente estudio y a la Estación Experimental Agraria (EEA) Los Cedros por el acceso a estereoscopios y microscopios electrónicos empleados en el fotoregistro de nematodos.
REFERENCIAS BIBLIOGRÁFICAS
Andree,
K. B., Loi, B., Vallainc, D., Concu, D., Duncan, N.,
& Carboni, S. (2024). Investigation of the utility of PCR-RFLP as a rapid
alternative to DNA sequencing for interrogation of the genetic sex of Mugil
cephalus. Animal Reproduction Science 270, 107614. https://doi.org/10.1016/j.anireprosci.2024.107614
Bhat, K. A., Mir, R. A., Farooq, A., Manzoor, M., Hami, A., Allie, K. A., ... & Shah, A. A. (2022). Advances in nematode identification: A journey from fundamentals to evolutionary aspects. Diversity, 14(7), 536. https://doi.org/10.3390/d14070536
Bikandi, J., Millán, R. S., Rementeria, A., & Garaizar, J. (2004). In silico analysis of complete bacterial genomes: PCR, AFLP–PCR and endonuclease restriction. Bioinformatics, 20(5), 798-799. https://doi.org/10.1093/bioinformatics/btg491
Bogale, M., Baniya, A., & DiGennaro, P. (2020). Nematode identification techniques and recent advances. Plants, 9(10), 1260. https://doi.org/10.3390/plants9101260
Campos-Herrera,
R., Escuer, M., Robertson, L., & Gutiérrez, C.
(2006). Morphological and ecological characterization of Steinernema feltiae
(Rhabditida: Steinernematidae) Rioja strain isolated from Bibio hortulanus
(Diptera: Bibionidae) in Spain. Journal of Nematology 38(1), 68.
Carta,
L. K., & Li, S. (2018). Improved 18S small subunit rDNA
primers for problematic nematode amplification. Journal of Nematology 50(4),
533. https://doi.org/10.21307/jofnem-2018-051
Chaves-Velasquez,
Á. M., Guzmán-Piedrahita, Ó. A., Valencia-Jíménez, A., & Villegas-Estrada,
B. (2023).
Caracterización
morfométrica y molecular de poblaciones nde Radopholus similis [(Cobb)
Thorne (Rhabditida: Pratylenchidae)] en Colombia. Boletín Científico. Centro
de Museos. Museo de Historia Natural 27(2), 97-116.
https://doi.org/10.17151/bccm.2023.27.2.6
Chen, Y., Long, H., Feng, T., Pei, Y., Sun, Y., & Zhang, X. (2022). Development of a novel primer–TaqMan probe set for diagnosis and quantification of Meloidogyne enterolobii in soil using qPCR and droplet digital PCR assays. International Journal of Molecular Sciences 23(19), 11185. https://doi.org/10.3390/ijms231911185
Chena, L., Nara, E., Sánchez, Z., Espínola, E., & Russomando, G. (2014). Estandarización de la técnica PCR-RFLP del gen mitocondrial cyt b como herramienta para la identificación de fuentes de alimentación de insectos hematófagos. Memorias del Instituto de Investigaciones en Ciencias de la Salud 12(2), 1780.
Chowdhury, I. A., & Yan, G. (2021). Development of real-time and conventional PCR assays for identifying a newly named species of root-lesion nematode (Pratylenchus dakotaensis) on soybean. International Journal of Molecular Sciences 22(11), 5872. https://doi.org/10.3390/ijms22115872
Córdoba-Sánchez, M., Rubiales, D., Cantalapiedra-Navarrete, C., Castillo, P., & Palomares-Rius, J. E. (2026). Plant-parasitic nematode diversity in Spanish pea fields with species-specific qPCR-based identification of Pratylenchus thornei and Pratylenchus mediterraneus. Plant Disease (ja). https://doi.org/10.1094/PDIS-09-25-1944-RE
Córdova-Campos, J., Calle-Ulfe, P. G., Suarez-Peña, E., Mendez-Farroñan, S., Lindo-Seminario, D., Gutiérrez-Calle, S., Morales-Pizarro, A., Cedeño-Escobar, V., Mialhe-Matonnier, E., & Condemarín-Montealegre, C. (2023). Identification of effecting proteins by MALDI TOF/TOF mass spectrometry of the root-knot nematode Meloidogyne javanica. Tropical and Subtropical Agroecosystems 26, 026. http://doi.org/10.56369/tsaes.4518
Córdova Campos, J. S., Riojas-González, J. M., Castillo-Carrillo, P. S., Cornejo-Hidalgo, R. E., Mogollón-Farias, C. A., Garcia-Garcia, S. M., Rosillo-Urbina, E. S., Ruiz-Turpo, Y. E., & Ruiz-Polo, A. A. (2025). Caracterización molecular y filogenética del gen 18S ARNr del nemátodo entomopatógeno Heterorhabditis indica aislado en el norte de Perú. Manglar 22(2), 203-209. https://doi.org/10.57188/manglar.2025.022
Fang, W., Xu, S., Zhang, S., Wang, Y., Chen, X., & Luo, D. (2010). Multiple primer PCR for the identification of anisakid nematodes from Taiwan Strait. Experimental Parasitology 124(2), 197-201. https://doi.org/10.1016/j.exppara.2009.09.011
Floyd, R., Abebe, E., Papert, A., & Blaxter, M. (2002). Molecular barcodes for soil nematode identification. Molecular Ecology 11(4), 839-850. https://doi.org/10.1046/j.1365-294X.2002.01485.x
Floyd, R. M., Rogers, A. D., Lambshead, P. J. D., & Smith, C. R. (2005). Nematode‐specific PCR primers for the 18S small subunit rRNA gene. Molecular Ecology Notes 5(3), 611-612. https://doi.org/10.1111/j.1471-8286.2005.01009.x
Gao, M., Zhang, M., Zhang, J., Yang, X., Abdallah, M. F., & Wang, J. (2023). Phylogenetic variation of Tri1 gene and development of PCR–RFLP analysis for the identification of NX Genotypes in Fusarium graminearum Species Complex. Toxins 15(12), 692. https://doi.org/10.3390/toxins15120692
Gaugler, R. (Ed.). (2002). Nematología entomopatógena. CABI Publishing.
Goraya, M., & Yan, G. (2024). Rapid and direct detection of the stubby root nematode Paratrichodorus allius from soil DNA extracts using recombinase polymerase amplification assay. International Journal of Molecular Sciences 25(19), 10371. https://doi.org/10.3390/ijms251910371
Hodson, A. K., Celayir, T., & Quiroz Alonso, A. (2023). A real-time PCR assay to detect and quantify root-knot nematodes from soil extracts. Plant Disease 107(7), 2169-2176. https://doi.org/10.1094/PDIS-10-22-2431-RE
Hu, M. X., Zhuo, K., & Liao, J. L. (2011). Multiplex PCR for the simultaneous identification and detection of Meloidogyne incognita, M. enterolobii, and M. javanica using DNA extracted directly from individual galls. Phytopathology 101(11), 1270-1277. https://doi.org/10.1094/PHYTO-04-11-0095
Huang, D., Yan, G., & Skantar, A. M. (2017). Development of real-time and conventional PCR assays for identifying stubby root nematode Paratrichodorus allius. Plant Disease 101(6), 964-972. https://doi.org/10.1094/PDIS-10-16-1431-RE
Huang, D., Yan, G., Gudmestad, N., Ye, W., Whitworth, J., Frost, K., ... & Hajihassani, A. (2019). Developing a one-step multiplex PCR assay for rapid detection of four stubby-root nematode species, Paratrichodorus allius, P. minor, P. porosus, and Trichodorus obtusus. Plant Disease 103(3), 404-410. https://doi.org/10.1094/PDIS-06-18-0983-RE
Huang, W. K., Heng, J. I. A. N., Abdulsalam, S., & Huan, P. E. N. G. (2023). Molecular diagnosis and direct quantification of cereal cyst nematode (Heterodera filipjevi) from field soil using TaqMan real-time PCR. Journal of Integrative Agriculture 22(8), 2591-2601. https://doi.org/10.1016/j.jia.2022.09.016
Inglehart, J., & Nelson, P. C. (1994). On the limitations of automated restriction mapping. Bioinformatics 10(3), 249-261. https://doi.org/10.1093/bioinformatics/10.3.249
Koppenhöfer, A. M., Shapiro-Ilan, D. I., & Hiltpold, I. (2020). Entomopathogenic nematodes in sustainable food production. Frontiers in Sustainable Food Systems, 4, 125. https://doi.org/10.3389/fsufs.2020.00125
Lacey, L. A., Frutos, R., Kaya, H. K., & Vails, P. (2001). Insect pathogens as biocontrol agents: do they have a future? Biological Control 21(3), 230-248. https://doi.org/10.1006/bcon.2001.0938
Laref, N., Belkheir, K., Belazreg, M., & Hireche, A. (2024). RFLP-kenzy: a new bioinformatics tool for in silico detection of key restriction enzyme in RFLP technique. Beni-Suef University Journal of Basic and Applied Sciences, 13(1), 83. https://doi.org/10.1186/s43088-024-00531-8
Leonar, A. L., Nimkingrat, P., Aryal, S., Martinez, J. G., Bhat, A. H., & Sumaya, N. H. (2022). Natural association of the entomopathogenic nematode Heterorhabditis indica (Rhabditida: Heterorhabditidae) from the Philippines with the non-symbiotic bacterium Ochrobactrum anthropi (Proteobacteria: Brucellaceae). Egyptian Journal of Biological Pest Control 32(1), 1-11. https://doi.org/10.1186/s41938-022-00576-w
Madikadike, M. K., Tyasi, T. L., Mbazima, V., & Gunya, B. (2024). Genetic polymorphism of growth hormone gene in Dorper sheep using PCR-RFLP. Indian Journal of Animal Research 58(3), 371-375. https://doi.org/10.18805/IJAR.BF-1683
Montero, Z., García, C., Salazar, L., Valverde, R., & Gómez-Alpízar, L. (2007). Detección de Meloidogyne incognita en tubérculos de papa en Costa Rica. Agronomía Costarricense 31(1), 77-84. https://doi.org/10.15517/rac.v31i1.6822
Niloufar, M. (2020). Diagnosis of potato rot nematode Ditylenchus destructor using PCR-RFLP. Вестник Российского университета дружбы народов. Серия: Агрономия и животноводство, 15(4), 353-362. https://doi.org/10.22363/2312-797X-2020-15-4-353-362
Nurashikin-Khairuddin, W., Abdul-Hamid, S. N. A., Mansor, M. S., Bharudin, I., Othman, Z., & Jalinas, J. (2022). A review of entomopathogenic nematodes as a biological control agent for red palm weevil, Rhynchophorus ferrugineus (Coleoptera: Curculionidae). Insects, 13(3), 245. https://doi.org/10.3390/insects13030245
Orui, Y. (1998). Identification of Japanese species of the genus Meloidogyne (Nematoda: Meloidogynidae) by PCR-RFLP analysis. Applied entomology and zoology, 33(1), 43-51. https://doi.org/10.1303/aez.33.43
Peraza-Padilla, W. (2021). Identificación morfológica y molecular de Meloidogyne javanica en una plantación de papaya (Carica papaya L.) en Pococí, Limón, Costa Rica. Agronomía Costarricense 45(2), 41-55. http://dx.doi.org/10.15517/rac.v45i2.47766
Perteguer, M. J., Ortiz, G., García, E., Flores, M., Rodríguez, E., Ubeira, F. M., & Gárate, T. (2004). Aplicación de la técnica PCR-RFLP en la identificación de nematodos implicados en la anisakiasis humana. Medicina Clínica 122(18), 686-689. https://doi.org/10.1016/S0025-7753(04)74356-0
Peteira, B., Rodríguez, M. G., Rosales, C., Maselli, A., Casado, R., Castro, L., Salazar, E., Enrique, R., & Miranda, I. (2014). Variabilidad molecular de aislamientos venezolanos de nematodos entomopatógenos y sus bacterias simbiontes. Revista de Protección Vegetal 29(2), 112-121.
Peraza-Padilla, W., Rosales-Flores, J., Esquivel-Hernández, A., Hilje-Rodríguez, I., Molina-Bravo, R., & Castillo-Castillo, P. (2013). Identificación morfológica, morfométrica y molecular de Meloidogyne incognita en higuera (Ficus carica L.) en Costa Rica. Agronomía Mesoamericana 24(2), 337-346. https://doi.org/10.15517/am.v24i2.12533
Poinar, G. O. (1979). Nematodes for biological control of insects (1st ed.). CRC Press. https://doi.org/10.1201/9781351074957
Půža, V., & Tarasco, E. (2023). Interactions between entomopathogenic fungi and entomopathogenic nematodes. Microorganisms 11(1), 163. https://doi.org/10.3390/microorganisms11010163
Rangarajan, V., Shahoveisi, F., Waldo, B. D., & Jafari, S. (2025). Identification of plant-parasitic nematode genera in turfgrass using deep learning algorithms. Scientific Reports, 16, 24. https://doi.org/10.1038/s41598-025-29467-4
Ren, Y., Porazinska, D. L., Ma, Q., Liu, S., Li, H., & Qing, X. (2024). Un único cebador degenerado mejora significativamente el rendimiento de la codificación de barras COX1 en la caracterización de la comunidad de nematodos del suelo. Soil Ecology Letters 6(2), 230204. https://doi.org/10.1007/s42832-023-0204-4
Rodríguez Zamora, M. J. (2019). Caracterización de aislados nativos de nematodos entomopatógenos y uso potencial contra Spodoptera frugiperda [Tesis de maestría, Universidad Nacional Agraria]. Repositorio digital de la Universidad Nacional Agraria.
Ruiz-Polo, A. A., Santillan-Valdivia, R. E., Saavedra-Rios, C. Y., Nuñez-Rodriguez, C. M., & Niño-Mendoza, L. E. (2024). Comportamiento alimentario de Aedes aegypti en brotes de dengue de dos zonas rurales del Perú durante el ciclón Yaku y El Niño Global del 2023. Revista Peruana de Medicina Experimental y Salud Pública 41(3), 266-272. https://doi.org/10.17843/rpmesp.2024.413.13696
Ruiz Polo, A. A., Luis Arismendiz, L. D., Barrera Rivera, L. V., Alvarado Aldana, A., Saavedra Cornejo, K. I., & Juárez Vilchez, J. P. (2024). Coexistencia y fuentes de alimentación de mosquitos adultos (Diptera: Culicidae) en un centro de salud rural en Piura, Perú 2024. Revista Peruana de Medicina Experimental y Salud Pública 41(3), 309-315. https://doi.org/10.17843/rpmesp.2024.413.13696
Rusinque, L., Camacho, M. J., Serra, C., Nobrega, F., & Inacio, M. L. (2023). Root-knot nematode assessment: species identification, distribution, and new host records in Portugal. Frontiers in Plant Science, 14, 1230968. https://doi.org/10.3389/fpls.2023.1230968
Sánchez Jara, J., Valle Delgado, J., Pérez Tesén, E., Neira de Perales, M., & Calderón Arias, C. (2019). Control biológico de Spodoptera frugiperda en cultivo de Zea mays: Uso de nematodos entomopatógenos. Scientia Agropecuaria 10(4), 551-557. https://doi.org/10.17268/sci.agropecu.2019.04.12
Shinde, S. P., Ingole, D. B., Biradar, V. K., Gokte-Narkhedkar, N., Lavhe, N. V., Thube, S. H., ... & Prasad, Y. G. (2022). Efficacy of native strains of entomopathogenic nematode Heterorhabditi indica against the fall armyworm, Spodoptera frugiperda (JE Smith) (Lepidoptera: Noctuidae) from India. Egyptian Journal of Biological Pest Control 32(1), 141. https://doi.org/10.1186/s41938-022-00638-z
Solano-González, S., Esquivel, A., Molina, R., & Morera, B. (2011). Identificación molecular de especies de nematodos noduladores (Meloidogyne spp.) asociadas a plantas ornamentales de altura. [Resumen de presentación en conferencia]. 2do Congreso Latinoamericano de Genética Humana, 6to Congreso Nacional de Biología, 2do Congreso Costarricense de Biotecnología, San José, Costa Rica.
Susurluk, A., Tarasco, E., Ehlers, R. U., & Triggiani, O. (2007). Molecular characterisation of entomopathogenic nematodes isolated in Italy by PCR-RFLP analysis of the ITS region of the ribosomal DNA repeat unit. Nematologia Mediterranea 35(1), 86913.
Torchia, J., Muzulín, P., & Martin, M. L. (2021). Identificación del reservorio de Fiebre Hemorrágica Argentina mediante la técnica de PCR-RFLP. Brazilian Journal of Animal and Environmental Research 4(1), 1329-1344. https://doi.org/10.34188/bjaerv4n1-109
Valderrama-Aravena, N., Pérez-Araneda, K., Avaria-Llautureo, J., Hernández, C. E., Lee, M., & Brante, A. (2014). Diversidad de nematodos marinos de Chile continental y antártico: una evaluación morfológica y molecular. Revista de Biología Marina y Oceanografía 49(1), 147-155. http://dx.doi.org/10.4067/S0718-19572014000100017
Vélez Zambrano, S. M., & Guzmán Cedeño, A. M. (2022). Técnicas de identificación del nematodo agallador Meloidogyne. Manglar 19(2), 209-215. https://doi.org/10.17268/manglar.2022.026
Vera-Morales, M., Quevedo, A., Peñaherrera, S., Serrano, L., Sosa, D., Ratti, M. F., ... & Magdama, F. (2024). Dinámica de crecimiento, esporulación y captura de hongos asexuales sobre Meloidogyne sp., in vitro. Scientia Agropecuaria 15(1), 121-132. http://dx.doi.org/10.17268/sci.agropecu.2024.009
Wang, W. H., Lin, C. Y., Jain, S. H., Lu, P. L., & Chen, Y. H. (2025). Development of the novel gene chip and restriction fragment length polymorphism (RFLP) methods for rapid detection of Mycobacterium tuberculosis complex in broth culture. Journal of Microbiology, Immunology and Infection 58(1), 56-61. https://doi.org/10.1016/j.jmii.2024.09.003
Yangüéz, J., Pitti, J., Vargas, R., Bado, M., Guerra, I., Martínez, O., & Ríos-Moreno, A. (2024). Patogenicidad de nematodos entomopatógenos sobre larvas de Agrotis sp. (Lepidoptera: Noctuidae). Revista Investigaciones Agropecuarias 7(1), 75-87. https://doi.org/10.48204/j.ia.v7n1.a6547
Ye, W., Robbins, R. T., & Kirkpatrick, T. (2019). Molecular characterization of root-knot nematodes (Meloidogyne spp.) from Arkansas, USA. Scientific reports, 9(1), 15680. https://doi.org/10.1038/s41598-019-52118-4
Ye, W., Koenning, S. R., Zeng, Y., Zhuo, K., & Liao, J. (2021). Molecular characterization of an emerging root-knot nematode Meloidogyne enterolobii in North Carolina, USA. Plant Disease, 105(4), 819-831. https://doi.org/10.1094/PDIS-04-20-0816-RE